




Жизнь растений
Основные систематические группы бактерий и актиномицетов (2)
Цитологические особенности спорообразующих анаэробов
Морфология и тонкое строение клеток анаэробных бактерий изучены еще слабо. Скудность морфологии клостридиев отмечалась многими микробиологами, изучавшими эту группу микроорганизмов. Лишь в последнее время с помощью фазово-контрастной и главным образом электронной микроскопии получены новые данные по цитологии анаэробов.
Принципиальных отличий в строении клеток анаэробов (в сравнении с аэробными организмами) не найдено. Однако некоторые группы спорообразующих анаэробов имеют определенные цитологические особенности. Так, анаэробные клостридии в отличие от аэробных споро-образующих бактерий не содержат липидных включений — гранул полибетагидроксибутирата (Имшенецкий, 1944). Вместо них у клостридиев и некоторых плектридиев накапливается гранулеза (крахмалоподобное соединение). И еще одной особенностью анаэробов, о которой упоминалось выше, является формирование на спорах анаэробов специфических выростов и ячеистых колпачков.
Физиологические и биохимические особенности спорообразующих анаэробных бактерий
Главные отличия анаэробов касаются энергетического метаболизма, т. е. реакций, снабжающих организм энергией, и ферментов, связанных с этими реакциями. Сущность анаэробиоза была кратко и в то же время точно сформулирована Луи Пастером как «жизнь в отсутствие воздуха», т. е. жизнь без дыхания, существующая за счет анаэробного превращения веществ.
Анаэробиоз свойствен и факультативно анаэробным микроорганизмам. В отличие от последних облигатные анаэробы не могут развиваться в присутствии кислорода, более того, кислород в молекулярной форме ядовит для анаэробов.
Как известно, микроорганизмы черпают энергию, необходимую для поддержания их жизнедеятельности, за счет различного рода процессов окисления органических (а иногда и неорганических) веществ. При этом окисление происходит путем отщепления от субстратов водорода (или электронов). Водород переносится по цепи ферментов и в конечном итоге соединяется с кислородом, образуя воду. Анаэробный же способ извлечения энергии характеризуется тем, что свободный кислород в нем не принимает участия, а органические субстраты окисляются только за счет отщепления водорода. Освобождающийся водород либо присоединяется к продуктам распада того же самого органического вещества, либо выделяется в газообразном состоянии.
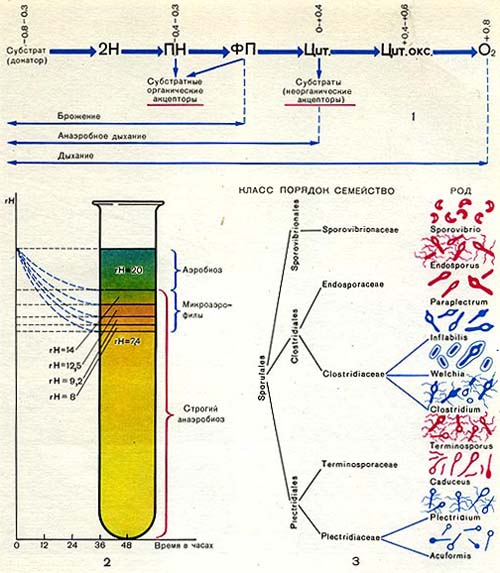
Таблица 36. Спорообразующие анаэробные бактерии.
На схеме 1 (табл. 36) показан транспорт электронов при дыхании и различных типах анаэробного способа получения энергии. Водород и электроны отщепляются от субстратов с помощью пиридиннуклеотидных ферментов (ПН). Далее они у аэробов проходят по цепи ферментов с возрастающими потенциалами — флавопротеидные (ФП)—цитохромные ферменты (Цит.) - и с помощью цитохромоксидазы (Цит. оке.) переносятся на кислород. Поток электронов направлен от системы с более низким (более отрицательным потенциалом) к системе с более высоким (более положительным) потенциалом, от — 0,8 — 0,4 в (потенциал субстрата) до + 0,8 в (потенциал кислорода).
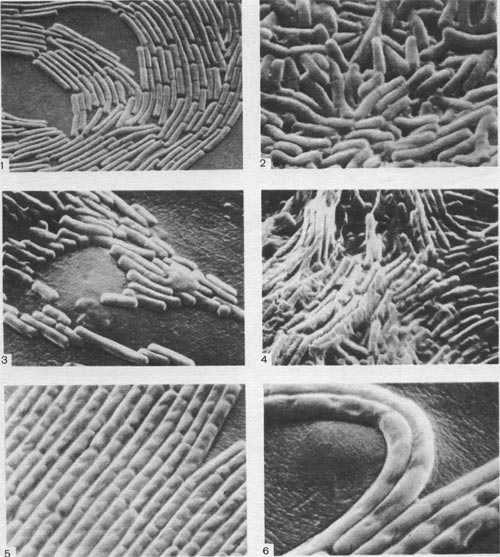
Таблица 37. Расположение клеток в колониях спорообразующих бактерий на поверхности плотных сред: 1—край колонии Bacillus cereus; 2 — центральная часть колонии Вас. cercus; 3 — край колонии Вас. mesentericus; 4 — центральная часть колонии Вас. mesentericus; 5, 6 — тяжи из клеток, образуемые Вас. mycoides. Фотографии сделаны Э. Г. Африкяном в сканирующем электронном микроскопе.
Таким образом, при дыхании конечным акцептором водорода является кислород. У анаэробов в качестве акцепторов водорода выступают либо органические субстраты (брожение), либо неорганические вещества, такие, как нитраты или сульфаты («анаэробное дыхание»). Из схемы видно, что наиболее просто и примитивно транспорт электронов осуществляется у большинства анаэробов из-за отсутствия у них ферментов цепи переноса электронов, способных передавать электроны по цепочке вплоть до молекулярного кислорода.
Молекулярный кислород отрицательно действует на рост и активность облигатных анаэробов. В присутствии свободного кислорода клетки анаэробов теряют подвижность. Благодаря этой реакции анаэробы были впервые обнаружены Пастером. Однажды он рассматривал под микроскопом каплю бродящей жидкости (при маслянокислом брожении), помещенную между двумя тонкими плоскими стеклышками, и заметил, что клетки, находящиеся по краям препарата (куда кислород воздуха . свободно проникал), становились внезапно неподвижными, а палочки, находящиеся в центре препарата (куда воздух не проникал), продолжали очень активно двигаться.
Отсюда Пастер сделал вывод, что кислород воздуха ядовит для некоторых микробов, и разделил последние на две группы — аэробов и анаэробов.
Противники Пастера (например, Трекюль) возражали против утверждения о существовании бактерий, для которых кислород воздуха может быть смертельным, и приводили в пример споры анаэробов, способные длительное время сохраняться на воздухе. На это Пастер отвечал, что споры не являются настоящими живыми существами, так как они не питаются и не размножаются. Последующее развитие науки подтвердило положения Пастера. Так, было показано, что спорам бактерий свойствен крайне выраженный анабиоз и обмен веществ у них находится на таком низком уровне, что его даже не удается измерить с необходимой точностью. В связи с этим споры не чувствительны ко многим повреждающим факторам, а споры анаэробов могут, кроме того (в отличие от вегетативных клеток), легко сохраняться на воздухе.
Вопрос, почему кислород является токсичным для анаэробов, еще недостаточно выяснен. Одни исследователи считают, что токсическое действие кислорода связано с образованием в культурах анаэробных микроорганизмов ядовитых концентраций перекиси водорода, образующейся в результате окисления субстрата кислородом воздуха. Причиной накопления перекиси водорода является отсутствие у анаэробов фермента каталазы (разлагающего перекись). Имеются сообщения, что некоторые штаммы клостридиев могут расти и в аэробных условиях, если к среде добавлена каталаза. Но, с другой стороны, очень важным фактором, определяющим развитие анаэробов, являются окислительно-восстановительные условия среды. Они выражаются через окислительно-восстановительный потенциал (ОВП), измеряемый в вольтах (напряжение на электроде, погруженном в испытуемую среду). Окислительно-восстановительные условия можно выразить также через показатель гНз, характеризующий соотношение между На и 02. rH2 в пределах от 0 до 40 характеризует все степени восстановленности или окисленности среды в зависимости от насыщения ее кислородом либо водородом. Подробные исследования условий развития анаэробов показали, что облигатные анаэробы не могут развиваться при rH2 (или ОВП) выше определенного предела. rНз в среде можно измерить либо электрометрическим способом . (с помощью потенциометров), либо с помощью красителей, которые восстанавливаются и обесцвечиваются (или изменяют цвет) при определенных значениях гНз. Так, например, краситель янусгрюн в аэробных условиях при rH = 20 и выше имеет в растворе зеленый цвет, при rН2 в пределах 12—14 — розовый цвет, а при еще более низких значениях rH он обесцвечивается.
А.Р. Прево проделал следующий опыт (схема 2 на табл. 36). Питательную среду (мясо-пептонный агар или желатин) он наливал в пробирки, кипятил 35 мин и оставлял на воздухе двое суток. Сразу после кипячения среда даже у самой поверхности была сильно восстановленной с низким гН2. Через 48 ч вследствие диффузии кислорода в среду показания гШ сильно изменились. С самой поверхности и до глубины 10 мм гШ составлял 20 единиц (янусгрюн — зеленый). Ниже 10 мм гНа был равен 14 (янусгрюн — розовый). На глубине 13 мм -12,5, при 16 мм — 9,2, а при 17 мм — 8, при 18 мм всего 7,4. Оказалось, что строгие анаэробы могут развиваться в средах с rH не выше 14.
Имеются также виды с высокой степенью анаэробности, которые начинают размножаться лишь при крайне низких значениях rH, приближающихся или почти равных нулю. В области rН2 выше 14 находится зона развития аэробов. В промежуточной области (между аэробиозом и строгим анаэробиозом) могут развиваться микроаэрофильные формы, приспособившиеся к развитию в средах с низкими концентрациями свободного кислорода.
При изучении действия кислорода на раз-. витие облигатных анаэробов было показано, что кислород не оказывает губительного действия на анаэробов, если ОВП среды низкий. Действительно, если к среде добавить восстанавливающие агенты, снижающие ОВП, то некоторые анаэробные микроорганизмы способны развиваться на таких средах в аэробных условиях. В целом анаэробы можно отнести к таким микроорганизмам, рост и развитие которых приурочены к природным субстратам, лишенным свободного кислорода и обладающим низким окислительно-восстановительным потенциалом.
Токсическое действие кислорода воздуха на рост и развитие облигатных анаэробов и тяготение к низкому окислительно-восстановительному потенциалу, по современным представлениям, можно объяснить тем, что молекулярный кислород и высокий ОВП могут служить причиной необратимого окисления жизненно важных ферментов, обусловливающих основные процессы их метаболизма.
Большинство анаэробных спорообразующих бактерий лишено также и других геминовых ферментов. Так, у бактерий из порядков Clostridiales и Plectridiales (по Прево) не обнаружены до настоящего времени цитохромы. Для переноса водорода в ряду субстрат — субстрат ими используются флавиновые ферменты, обладающие свойством аутооксидабильности (самоокисление кислородом воздуха). Хотя ферменты облигатных анаэробов (например, флавиновые) окисляются кислородом, он у них не может служить в качестве физиологического акцептора водорода. Реакция флавиновых ферментов с кислородом направлена на детоксикацию молекулярного кислорода.
Из спорообразующих анаэробов цитохромы (цитохромы бис) обнаружены только у суль-фатредуцирующих микроорганизмов рода De-sulfotomaculum. Эти бактерии также очень чувствительны к кислороду. Таким образом, различия между облигатными анаэробами и аэробами касаются прежде всего ферментативного обеспечения терминального окисления. У анаэробов свободный кислород не может быть использован в качестве конечного акцептора водорода.
В клетках спорообразующих анаэробов в энергетическом обмене веществ играют большую роль флавиновые ферменты.
Не удивительно поэтому, что содержание флавиновых дегидрогеназ в клетках анаэробов (маслянокислые бактерии) значительно выше, чем у аэробов.
Облигатные анаэробы представляют собой, очевидно, пример ранних анаэробных форм жизни. Это согласуется с теорией происхождения жизни на Земле, по которой первичными организмами нашей планеты были анаэробы. Сравнительный биохимический анализ приводит к выводу, что в основе энергетического обмена всех без исключения организмов лежат одни и те же поразительно сходные между собой цепи реакций, не связанных с потреблением свободного кислорода, — реакции, которые происходят в клетках современных анаэробов (по А. И. Опарину).
В связи с этим большое значение имеет также вопрос о локализации в клетках анаэробных микроорганизмов дегидрогеназ. Считалось, что у аэробов дегидрогеназы связаны с мембранами, а у анаэробов, не обладающих цитохрома-ми и окислительным фосфорилированием (кло-стридии), эти ферменты не связаны с мембранами и находятся в цитоплазме только в растворимой форме. Эта точка зрения как будто находится в согласии с данными о том, что ферменты гликолитического цикла (ферменты, участвующие в распаде углеводов в анаэробных условиях) находятся в клетках животных в растворимой форме.
Таким образом создавалось впечатление, что механизмы, обеспечивающие анаэробиоз, в структурном отношении организованы более примитивно. Свидетельством этого является отсутствие в данном случае связи дегидрогеназ с мембранами. Однако недавно было показано, что ферменты гликолитического цикла (в том числе дегидрогеназы) локализуются в плазматической мембране.
Ошибочность прежних данных объяснялась грубостью методов выделения мембран для биохимических анализов. Выделение чистых фракций мембран включает жесткие приемы разрушения клеток и последующее разделение осколков клеток на фракции различных клеточных структур. В процессе таких обработок ферменты, связанные с мембранами, могут легко от них отделиться. Как отмечают известные биохимики Д. Грин и Р. Гольдбергер (1968), именно эта хрупкость мембран вводила в заблуждение два поколения биохимиков, которые считали, что гликолити-ческие ферменты не связаны с мембранами.
Применение электронно-микроскопических методов, позволяющих изучать распределение дегидрогеназ в целых клетках, показывает, что дегидрогеназы у анаэробных спороносных бактерий, очевидно, также связаны с мембранами, играющими у живых организмов огромную роль, особенно в процессах энергетического метаболизма. Например, у такого облигатного анаэроба, как Cl. taeniosporum, активность окислительно-восстановительных ферментов проявлялась только в связи с цитоплазматиче-ской мембраной (рис. 60). В то же время у других анаэробов восстановление акцепторов электронов наблюдается и в цитоплазме. Возможно, эти явления связаны с различным набором ферментов у разных видов либо с неспецифическим восстановлением красителей в цитоплазме.
В качестве индикаторов для окислительно-восстановительных ферментов в электронной микроскопии используют теллурит калия или соли тетразолия. Эти соединения, будучи добавленными в среду и проникая в клетки, акцептируют электроны, которые они получают от дегидрогеназ. В результате реакции эти соединения восстанавливаются и выпадают в осадок, имеющий вид электронно-плотных (темных на экране микроскопа) зернышек, глыбок, тонких слоев. Теллурит в качестве индикатора окислительно-восстановительных ферментов более показателен, чем тетразолии, так как соли тетразолия могут неспецифически восстанавливаться веществами, содержащими сульфгид-рильные группы, а также редуцирующими (восстанавливающими) сахарами. Особенно это относится к анаэробным микроорганизмам, развивающимся при очень низком окислительно-восстановительном потенциале среды и вырабатывающим различные вещества с сильной редуцирующей активностью. Весьма интересно, что у некоторых анаэробных бактерий рода Clostridium обнаружена специфическая, но эффективная форма окислительного фосфорилиро-вания (фосфорилирования в цепи окислительных ферментов).
Если согласиться с утверждением (довольно убедительно обоснованным данными сравнительной физиологии и биохимии) о том, что облигатные анаэробы — это пример ранних форм жизни на Земле, то возникает вопрос, не отразились ли происхождение и эволюция анаэробов на составе и строении их ДНК — хранителе генетической информации. Сейчас уже хорошо известно, что дезоксирибонуклеиновым кислотам всего органического мира присущ единый план строения, а с другой стороны, имеются безграничные возможности вариаций состава и структуры этих соединений. Вполне логично думать, что возникновение ДНК в истории жизни на Земле было весьма важным и, вероятно, даже решающим фактором в дифференциации и обособлении новых групп и видов живых существ. Так как именно нуклеиновые кислоты непосредственно связаны с наследственностью и изменчивостью, то они должны являться материальной основой эволюционного процесса.
Специфичность нуклеиновых кислот для различных организмов обусловливается их химическим строением и может отражаться в первую очередь на количественном соотношении пуриновых и пиримидиновых оснований (на нуклеотидном составе) и на последовательности чередования нуклеотидов в цепи молекулы. Состав ДНК бактерий очень сильно варьирует от самого крайнего АТ-типа (преобладает пара — аденин и тимин) до ГЦ-типа (преобладают гуанин и цитозин). Если выразить соотношение оснований в виде молярных процентов Г + Ц, то колебания этого показателя лежат в пределах от 25 до 75%. Весьма показательно в этом отношении, что сложно организованные прокариотные организмы -актиномицеты — имеют самый высокий процент Г + Ц в ДНК, а примитивные анаэробные спорообразующие бактерии, у которых отсутствуют цитохромы, обладают крайним АТ-типом(до25% Г + Ц).Средиспорообразующих анаэробов также можно выявить определенные закономерности. Так, для гетеротрофных, не содержащих цитохромов бактерий (клостридии) колебания Г + Ц находятся в пределах от 25 до 39%, а для спорообразующих сульфатредуцирующих анаэробов, являющихся хемолито-гетеротрофами и обладающих уже отдельными цитохромами, процент Г + Ц находится в пределах 45—48.
Таким образом, ДНК у анаэробов имеет интересные особенности, и очень многого можно ожидать в будущем от изучения первичной структуры ДНК этих бактерий.
Физиологические группы анаэробных спороносных бактерий
Облигатные анаэробные бактерии, способные образовывать эндогенные споры, в настоящее время можно подразделить в зависимости от характера используемых ими субстратов и особенностей метаболизма на следующие физиологические группы: сахаролитические, протеоли-тические, пуринолитические и сульфатредуцирующие анаэробные бактерии.
Сахаролитические анаэробные бактерии способны сбраживать различные углеводы (органические кислоты и спирты, полисахариды, пектиновые вещества). Обычно конечными продуктами брожений являются органические кислоты, растворители (спирт, ацетон, бутанол и др.), углекислый газ и водород. Было установлено существование нескольких типов брожения.
Маслянокислое брожение осуществляют такие анаэробы, как Cl. butyricum, Cl. tyrobu-tyricum, Cl. lactoacetophilum, и другие виды. Главные продукты брожения — масляная и уксусная кислоты. Так, сбраживание глюкозы Cl. butyricum можно описать уравнением
4 глюкоза —> 2 уксусная кислота + 3 масляная кислота + 8С02+8Н2
Образование масляной кислоты происходит также, если в среду вместо углеводов вносить молочную кислоту вместе с уксусной. Уксусная кислота играет роль акцептора водорода. Образовавшиеся при маслянокислом брожении кислоты далее микроорганизмом не потребляются.
Ацетонобутиловое брожение представляет собой биохимически более сложный тип масля-нокислого брожения, в котором образовавшиеся на первой стадии жирные кислоты превращаются в нейтральные конечные продукты -ацетон, бутиловый и изопропиловый спирты. Если в первой фазе брожения увеличивается кислотность, то во второй фазе кислотность не только не возрастает, но даже уменьшается. Таким образом, за счет превращения кислот в нейтральные продукты регулируется рН среды (кислотность). Ацетонобутиловое брожение углеводов вызывается Cl. acetobutylicum и близкими к нему по физиологии видами.
Cl. butylicum осуществляет брожение, очень сходное по типу с ацетонобутиловым, но отличается тем, что вместо ацетона в этом случае образуется изопропиловый спирт.
Уксуснокислое брожение
Уксуснокислое брожение углеводов наблюдается у Cl. aceticum и Gl. thermoaceticum. Уксусная кислота — единственный продукт брожения.
Пропионовокислое брожение
Пропионовокислое брожение присуще С1. propionicum. В качестве основных продуктов образуются пропионовая и уксусная кислоты, а также углекислый газ.
Брожение пектиновых веществ
Брожение пектиновых веществ (полиурониды) способны осуществлять анаэробные бактерии из различных систематических групп. В настоящее время известно около 18 видов кло-стридиев, способных разрушать пектиновые вещества. Наиболее активные пектинолити-ческие виды — Gl. felsineum, Cl. laniganii, Cl. pectinolyticum, Cl. pectinovorum, Cl. virens и другие пигментированные и непигментированные клостридии и плектридии. Имеются и факультативно-анаэробные формы — Cl. таcerans и Cl. polymyxa. Каждому из этих видов свойственны свои специфические детали обмена веществ, но общим их свойством является способность разлагать пектиновые вещества до органических кислот, спиртов и газов. Некоторые бактерии обладают очень стойкими пектинолитическими свойствами и выделяют пектинолитические ферментш на средах без пектиновых веществ. У других анаэробов (например, у Cl. multifermentans) синтез ферментов происходит только при добавлении в среды пектинов (индуцированный синтез ферментов).
Сбраживание сахаров пектинолитические анаэробы осуществляют по маслянокислому или ацетонобутиловому типу.
Брожение целлюлозы
Большая группа высокоспециализированных анаэробных спорообразующих бактерий получает энергию за счет сбраживания целлюлозы. Конечными продуктами брожения являются уксусная, пропионовая, масляная и молочная кислоты, этиловый спирт, водород и углекислый газ. В качестве промежуточных продуктов при гидролизе целлюлозы образуются глюкоза и целлобиоза. И весьма удивительно, что если добавить к среде глюкозу (или другие сахара), то они очень плохо усваиваются либо совсем не используются. А если в среде имеются одновременно глюкоза и клетчатка, то сбраживается прежде всего клетчатка. Это свидетельствует о высокой специализации целлюлозоразлагаю-щих анаэробов.Целлюлозолитические бактерии отличаются не только физиологическими, но и морфологическими особенностями. Большинство целлюлозолитических спорообразующих анаэробов имеют вид очень тонких длинных палочек, образующих споры по плектридиальному типу.
Обычно вегетативные клетки находятся в адсорбированном состоянии на волокнах целлюлозы (рис. 51). Вероятно, это связано с тем, что ферменты, гидролизующие клетчатку (целлюлазы), в среду не выделяются, а прикреплены к поверхности клеток. Под микроскопом можно увидеть, что разрушаются те участки клетчатки, к которым клетки непосредственно прикасаются. Спорообразующие клетки обычно находятся в растворе, так как при спорообразовании изменяется характер связи со средой, а спорообразование идет за счет эндогенного метаболизма (за счет внутриклеточных запасов питательных веществ).
Первая культура анаэробных спорообразующих бактерий, способная осуществлять брожение целлюлозы, была выделена В. Л. Омелянским. В литературе описано 12 видов целлюлозолитических анаэробов. Спорообразующие Целлюлозолитические анаэробы до настоящего времени очень плохо изучены в связи с трудностями их выделения и культивирования. Большинство выделенных штаммов были загрязнены посторонними бактериями, что значительно обесценивает данные по физиологии и биохимии этих организмов.
Брожение органических кислот и спиртов. Выделено несколько специализированных видов анаэробных бактерий (возможно, их в природе существует намного больше), использующих органические кислоты и спирты в качестве источника углерода и энергии.
Так, Cl. kluyveri получает энергию путем сопряженного окисления-восстановления системы этиловый спирт — уксусная кислота. В результате реакции образуются высшие жирные кислоты (главным образом капроновая и масляная кислоты). Характерно, что Cl. kluyveri не способен сбраживать углеводы, аминокислоты и пурины. Накопление энергии в форме АТФ происходит у этого анаэроба с использованием механизмов окислительного фосфорилирования. Cl. lactoacetophilum осуществляет брожение сходного типа, сбраживая смесь лак-тата и уксусной кислоты. Обе эти бактерии используют в качестве акцепторов водорода уксусную кислоту, а в качестве доноров первая — этанол, вторая — лактат. Cl. glycollicum превращает этиленгликоль в смесь этилового спирта и уксусной кислоты:
Анаэробная бактерия из рода Clostridium (не определенная до вида) осуществляет крайне интересный тип брожения соединений с сульфониевой связью. Такие вещества могут рассматриваться как соединения, обладающие мак-роэргической связью, при разрыве которой выделяется большое количество энергии. Clostridium species (Штадтман, 1968) сбраживает диметилпропиотетин, используя его в качестве единственного источника углерода и энергии:
3(CH3)2S+-CH2CH2COO-+2H20 -> -> 2CH3CH2COO-+CH3COO- + 3(CH8)2S+CO2+H+ пропионовая кислота уксусная кислота диметилсульфид диметилпропиотетин
Протеолитические анаэробные бактерии - очень большая группа анаэробов, получающая энергию за счет анаэробного разрушения аминокислот. Многие из них являются строго протеолитическими организмами, неспособными сбраживать углеводы. Другие обладают слабовыраженной сахаролитической активностью. Имеются также активные протеолитические виды, способные осуществлять маслянокислое брожение Сахаров. Протеолитическая активность у некоторых анаэробов (Cl. histolyticum) настолько высока, что у зараженных этими микроорганизмами животных наблюдается как бы расплавление мышц, после чего остаются голые кости. Освобождающиеся после протео-лиза (разрушение белков протеолитическими ферментами) аминокислоты могут сбраживаться двумя путями. Cl. sporogenes, например, сбраживает пару аминокислот, причем одна из них окисляется, а другая восстанавливается (реакция Стикленда), т. е. одна аминокислота служит донором, а другая — акцептором водорода:
аланин + 2 глицин + 2Н20 -> -> 3 уксусная кислота + C02+3NH3
Существуют виды протеолитических анаэробов, которые могут сбраживать и по одной аминокислоте, например глутаминовую кислоту (Cl. tetani).
Пуринолитические анаэробы
Существуют три вида бактерий, сбраживающих гетероциклические соединения с азотным кольцом: С1. acidiurici, Cl. cylindrosporum и Cl. uracilicum. Они способны разрушать пурины и пиримидины с образованием уксусной кислоты, углекислого газа и аммиака. Первые два вида не способны использовать ни углеводы, ни белки (аминокислоты). Ими быстро расщепляются ксантин, гуанин, гуанозин, 6,8-диоксипурин; более медленно и после адаптации (приспособления) разрушаются гипоксантин и инозин. Cl. acidiurici и G1. cylindrosporum физиологически очень близки, но четко разделяются по морфологическим признакам. У Cl. acidiurici споры имеют овальную форму и расположены терминально. Клетки при спорообразовании раздуваются. У Cl. cylindrosporum споры цилиндрические, расположены центрально или субтерминально. Спорангии при этом не раздуваются.
Сульфатредуцирующие бактерии. Восстановление микроорганизмами полностью или частично окисленных соединений минеральной серы в анаэробных условиях за счет окисления органического вещества или водорода называют микробной сульфатредукцией. Этот процесс был открыт русским химиком Н. Д. Зелинским в 1890—1893 гг. Чистая культура сульфатредуцирующих бактерий была выделена Бейеринком в 1895 г. Сначала бактерий, восстанавливающих сульфаты, относили к неспороносным. Затем было замечено, что в культурах сульфатредуцирующих бактерий иногда появляются споры (когда инкубирование проводилось при высоких температурах). В настоящее время выяснилось, что большинство неспорообразующих штаммов, выделенных из различных субстратов, было загрязнено спо-рообразующими. Последние выделены в новый род Desulfotomaculum, в который входят четыре вида: D. nigrificans, D. ruminis, D. orien-tis, D. antarcticum. Первые два вида используют некоторые органические кислоты (молочную, пировиноградную), но не усваивают углеводы (глюкозу). D. antarcticum способен усваивать глюкозу. D. nigrificans — термофил, оптимальная температура для роста этого вида 55° С. Восстановление сульфатов происходит по уравнению:
2СН3 - CHOH-COOH+Na,S04 -> - 2СН3 - COOH+2C02+H2S+NaOH молочная кислота сульфат натрия уксусная кислота
Таким образом, у анаэробных спорообразующих бактерий мы встречаемся с удивительным физиологическим разнообразием как по механизмам анаэробного получения энергии, так и по диапазону используемых субстратов.
Имеется группа анаэробов, использующих главным образом углеводы; другая группа разлагает преимущественно белки и аминокислоты; третья — сбраживает только пурины; четвертая — получает энергию за счет окисления органических кислот или водорода, используя в качестве акцептора водорода сульфаты.
Специфичность по отношению к субстратам у спорообразующих анаэробов выражена резко.
Потребности в питательных веществах у этих групп весьма различны. Некоторые протеолитические анаэробы (например, Cl. sporogens) не удовлетворяются средами, содержащими полный набор аминокислот, углеводы, минеральные соли, комплекс витаминов, активаторы микробного роста. Такие резко выраженные гетеротрофы растут лишь на средах, содержащих белки или продукты их частичного гидролиза. С другой стороны, существуют анаэробы, развивающиеся на очень простых средах, в состав которых входит несколько минеральных солей, в том числе сульфаты и органическая кислота либо водород в качестве донора электронов (сульфатредуцирующие бактерии). При этом может усваиваться атмосферный азот. Способность фиксировать молекулярный азот широко распространена среди спороносных бактерий. Этот процесс могут осуществлять маслянокислые, ацетонобутиловые и сульфатредуцирующие бактерии. Наиболее активные азотфиксаторы — сахаролитические анаэробы (клостридии).
Отношение к кислороду не одинаковое у различных физиологических групп спорообразующих анаэробов. Более устойчивыми к действию кислорода являются сахаролитические анаэробы. Часть представителей этой группы -аэротолерантные формы (способные давать слабый рост на пластинках агара даже в аэробных условиях), например Cl. carnis, Cl. histolyticum и др. Очень чувствительны к кислороду и трудно культивируются сульфатредуцирующие бактерии. Они могут развиваться только в строго анаэробных условиях при тщательном удалении кислорода из сред, в которых они культивируются.
Распространение и роль в природе спорообразующих анаэробных бактерий
Анаэробные спорообразующие бактерии широко распространены в природе. Их находили в самых различных почвах всех континентов, на скальных породах горных массивов, в морях, пресных водах озер и рек, водах глубинных нефтяных источников, в кишечнике человека и животных и т. д. В определителе А. Р. Прево дано описание около 200 видов анаэробных спорообразующих бактерий. На самом деле их в природе гораздо больше. Общепризнанным является мнение, что только меньшая доля микроорганизмов известна в настоящее время. Особенно это касается анаэробов, которые с большим трудом выделяются из природных субстратов и культивируются в лабораториях. И не случайно с каждым годом мы узнаем о новых микроорганизмах, выделенных «охотниками» за микробами.
С открытием новых, особенно своеобразных, видов и групп микроорганизмов становятся известными новые биохимические процессы, осуществляемые микроорганизмами в природе, новые метаболиты микроорганизмов (ферменты, гормоны, витамины, антибиотики и т. д.), не говоря уже о том, что расширяются наши представления о разнообразии форм микроорганизмов и органического мира в целом. Благодаря очень интенсивному обмену веществ, а следовательно, и необычайно высокой химической активности, микроорганизмы играют огромную роль в круговороте веществ на Земле. Анаэробные микроорганизмы принимают активное участие во многих звеньях этого круговорота, что обусловлено разнообразием и сложностью процессов их обмена веществ, большим набором органических и неорганических веществ, трансформируемых этими микроорганизмами, и крайне широким спектром выделяемых продуктов метаболизма. Во всех биологических циклах (круговороте углерода, азота, серы, фосфора и других элементов) анаэробам принадлежит весьма существенная роль.
Сахаролитические спороносные анаэробы «специализируются» на разложении углеводов и родственных им соединений, попадающих в почву и другие среды благодаря деятельности фотосинтезирующих организмов.
Другие физиологические группы анаэробов участвуют в круговороте азотсодержащих веществ: разлагают белки, аминокислоты, пурины (протеолитические, пуринолитические бактерии). Многие же способны активно фиксировать атмосферный азот, переводя его в органическую форму. Эти анаэробы способствуют повышению плодородия почв. Количество клеток протеолитических и сахаролитических анаэробов в 1 г плодородных почв достигает даже миллионов. Особое значение имеют те группы микроорганизмов, которые участвуют в разложении труднодоступных форм органических соединений, таких, как пектиновые вещества и целлюлоза. Именно эти вещества составляют большую долю растительных остатков и являются главным источником углерода для почвенных микроорганизмов. В анаэробных условиях эти соединения разрушаются пекти-нолитическими и целлюлозолитическими анаэробами.
Деятельность сульфатредуцирующих бактерий очень многогранна. С ними связывают образование нефти, сероводорода в морях, почвах, лечебных грязях. Еще в 1890 г. Н. Д. Зелинский и Е. Брусиловский показали, что слой воды в Черном море, насыщенный сероводородом, является результатом активности сульфатредуцирующих анаэробных бактерий. В почвах деятельность сульфатредуцирующих бактерий приводит к содообразованию. В определенных условиях, благоприятствующих накоплению соды, может произойти содовое засоление почвы. Массовое развитие сульфатредуцирующих бактерий, например, в почвах рисовых плантаций приводит к усилению восстановительных процессов, переводящих многие питательные вещества в доступную для корней риса форму.
Образование сероводорода сульфатредуци-рующими бактериями часто приносит вред, вызывая при определенных обстоятельствах коррозию металлических подводных и подземных сооружений.
Патогенные и токсигенные спорообразующие анаэробы
Некоторые из протеолитических и сахаролитических бактерий могут быть возбудителями болезней, особенно таких, как гангрена и столбняк (так называемые раневые инфекции). Возбудителями газовой гангрены являются такие виды анаэробных спороносных бактерий, как Cl. perfringens, Cl. histolyticum, Cl. septicum, Cl. oedematiens, Cl. bifermentans. Возбудитель столбняка — Cl. tetani. Хотя патогенные бактерии не так часто встречаются в медицинской практике, как другие патогенные микроорганизмы, но вызываемые ими заболевания очень опасны, протекают быстро и часто с фатальным исходом.
Попадая в рану, гангренозные формы анаэробов вызывают быстро распространяющееся разрушение мышечной ткани, сопровождающееся накоплением в ткани водорода. Возбудители вырабатывают сильнодействующие экзотоксины. Наиболее изучен альфа-токсин, выделяемый Cl. perfringens. Это фермент лецитиназа, разрушающая лецитин (важный компонент клеточных мембран). Лецитиназа также лизирует и красные кровяные тельца. Cl. tetani при развитии в ранах продуцирует белковый токсин большой силы, который через кровь и нервные каналы быстро проникает в организм и вызывает повреждение центральной нервной системы.
Cl. botulinum продуцирует токсины, вызывающие опасные, часто фатальные пищевые отравления. Насчитывается 5 разновидностей Cl. botulinum, синтезирующих 6 различных токсинов. Из них наибольшей токсичностью обладает токсин типа Д, 1 мг которого содержит около 1012 минимальных летальных доз для мышей; 5 —10 мг этого токсина способны убить все население земного шара. Это наиболее сильный из всех токсинов, которые способны вырабатывать живые организмы. При пищевых отравлениях чаще всего встречается токсин типа А. Несмотря на большую молекулярную массу (—1000000), токсин легко проникает через стенку кишечника в кровь. Смерть обычно наступает в результате остановки дыхания. Токсин очень устойчив к действию протеолитических ферментов кишечного тракта, но термолабилен и может разрушаться при кипячении.
Использование анаэробов в промышленности
Получение ацетона и бутанола с помощью микроорганизмов было первым крупнотоннажным производством, где нашли применение анаэробные спорообразующие бактерии. В качестве продуцента ацетона и бутанола использовался Clostridium acetobutylicum, вызывающий ацетонобутиловое брожение углеводов.
Ацетон и бутанол — ценное сырье для производства взрывчатых веществ и каучука. Бутиловый спирт — один из лучших растворителей целлюлозных лаков. Особенно широко начал применяться микробиологический способ получения растворителей в конце 40-х годов. В последние годы расширяется использование микроорганизмов как в нашей стране, так и за рубежом для промышленного производства таких ценных веществ, как антибиотики, ферменты, аминокислоты, гормоны, витамины, белкововитаминные препараты.
Дальнейший прогресс микробиологической промышленности связан с поисками и внедрением новых перспективных продуцентов веществ, необходимых для народного хозяйства. В этом отношении экономически выгодно может быть применение анаэробных микроорганизмов, в частности спорообразующих анаэробов, которые, как известно, являются активными продуцентами ряда важных ферментов: пектиназ, цел-люлаз, протеаз, липаз, амилаз, декарбоксилазы и гиалуронидазы. Анаэробы в некоторых странах уже используются для трансформации стероидных гормонов, а также для получения липазы, коллагеназы и пектаз. Так как при выращивании анаэробов воздух (кислород) в ферментеры подавать не нужно, производственный процесс облегчается, а стоимость продуктов ферментации значительно удешевляется по сравнению с аналогичными аэробными процессами.
В последнее время показано, что споры некоторых видов анаэробов при внутривенном введении животным проявляют онколитическую активность (наблюдается разрушение злокачественных образований). Споры анаэробов обладают также радиопротекторной активностью и при внутривенном введении животным способны повышать выживаемость и среднюю продолжительность жизни животных, облученных смертельной дозой радиации. Очевидно, споры анаэробов можно использовать для получения онколитических и радиопротекторных веществ.
Спороносные анаэробные бактерии применяются при мочке льна, конопли и других лубоволокнистых растений. Под влиянием микробных ферментов (пектиназ) расщепляются пектиновые вещества, склеивающие паренхим-ные клетки и лубяные пучки между собой. Этот процесс используется для получения волокна, необходимого для текстильной промышленности. До сих пор в промышленных условиях мочку льна осуществляют без регулирования состава пектинолитических микроорганизмов. В то же время применение в виде заквасок чистых культур анаэробов (активных пектинолитических клостридиев) сокращает процесс в 3—4 раза и позволяет получать волокно высокого качества. Применение специальных препаратов позволит в будущем, кроме того, стандартизовать процесс получения льняного волокна. Пектинолитические ферменты анаэробов имеют в этом отношении преимущество по сравнению с ферментами грибов, которые содержат примеси целлюлаз, ухудшающих качество волокна.
Дальнейшее всестороннее изучение обширного мира анаэробных бактерий не только приведет ко многим важным открытиям теоретического характера, но и будет способствовать широкому практическому использованию деятельности анаэробных микроорганизмов в народном хозяйстве.
Таблица 38. Нитевидные формы анаэробных бактерий, растущих на почвенных частицах (фото В. И. Дуды).
Таблица 39. Спорангии и споры у актиномицетов.
Систематика спорообразующих анаэробных бактерий
В настоящее время имеются три наиболее известные системы классификации анаэробов: Н. А. Красильникова (1949), Берги (1957) и Прево (1957, 1967). По Красильникову, все спорообразующие бактерии объединяются в семейство ВасШасеае. Это семейство имеет два рода — Bacillus и Clostridium. В род Bacillus входят как аэробные, так и анаэробные бактерии (последние составляют Х-группу), которые не образуют гранулезы и не раздуваются (либо слабо раздуваются) при спорообразовании. Род Clostridium объединяет бактерии, клетки которых в процессе спорообразования раздуваются с приобретением клостридиальной (лимоновидной) формы и накапливают гранулезу.
За прошедшие 20 лет после издания определителя Красильникова накопилось много данных об анаэробных микроорганизмах, их видовом разнообразии, биохимии, физиологии и цитологии. Были обнаружены новые физиологические группы, относящиеся к разным родам, и выделено много новых видов спорообразующих анаэробов.
Определитель Прево, Турпина и Кайзера наиболее полно описывает анаэробные микроорганизмы, известные в настоящее время микробиологам. Около 200 видов спорообразующих анаэробных бактерий распределяются в нем по трем порядкам: Clostridiales, Plectridiales и Sporovibrionales (схема 3 на табл. 36).
Морфологические различия между вегетативными клетками различных видов очень незначительны, а спорулирующие клетки и сами споры необычайно разнообразны. Это явление имеет очень большое значение для разделения анаэробов на систематические группы. Для разделения на порядки служит форма вегетативных клеток (прямые или изогнутые палочки) и расположение спор в спорангиях (центральное, субтерминальное и терминальное). При разделении на семейства и роды принимаются во внимание грамокраска, подвижность клеток, способность образовывать капсулу. Различные сочетания этих признаков позволили разделить все спорообразующие анаэробные бактерии на 5 семейств и 10 родов.
Порядок Clostridiales. К нему относятся подвижные и неподвижные грамположительные и грамотрицательные палочки, раздувающиеся при спорообразовании. Споры расположены центрально и субтерминально. Имеются пигментированные виды. В основном анаэробные, но возможны факультативные формы. В порядок входят два семейства.
Семейство Endosporaceae включает грамотрицательные клостридии; споры расположены центрально или субтерминально. В этом семействе два рода: Endosporus — подвижные формы и Paraplectrum — неподвижные, образуют капсулу. Семейство Clostridiaceae включает грамположительные клостридии. Споры центральные или субтерминальные. Подразделяется на три рода: Inflabilis — неподвижные формы, Welchia — неподвижные капсулированные палочки, Clostridium — подвижные формы.
Порядок Plectridiales. Подвижные и неподвижные, грамположительные и грамотрицательные палочки. Споры образуются терминально. Порядок подразделяется на два семейства.
Семейство Terminosporaceae — грамотрицательные плектридии. Семейство состоит из двух родов: Terminosporus — подвижные и Cadu-ceus — неподвижные формы.
Семейство Plectridiaceae — грамположительные плектридии. В этом семействе два рода: Plectridium — подвижные и Acuformis — неподвижные организмы.
Порядок Sporovibrionales включает спорообразующие подвижные анаэробные бактерии с изогнутыми (вибрионоподобными) грамотрицательными клетками. Большинство из них способно восстанавливать сульфаты и нитраты.
В этом порядке одно семейство — Sporovibrionaceae. В него входит один род — Sporovibrio, объединяющий четыре вида спорообразующих анаэробов. Один из этих видов — Sporovibrio ferrooxydans — не восстанавливает сульфатов, а железо окисляет в гидрат железа. По Кэмпбеллу и Постгейту, все спорообразующие анаэробы, обладающие постоянной способностью восстанавливать сульфаты, были выделены в новый род — Desulfo-tomaculum. В него входят облигатные анаэробы с грамотрицательными, прямыми или изогнутыми палочками, раздувающимися у термофильных форм. Споры формируются терминально или субтерминально. Бактерии подвижные, жгутикование перитрихиальное, восстанавливают сульфаты до сульфидов. Обладают цито-хромами (класс протогеминов). Состав ДНК колеблется в пределах 41,7—49,2 мол.% Г+Ц.
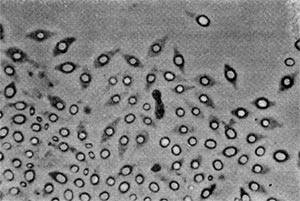
Рис. 64. Сульфатредуцирующая спороносная бактерия. Клетки при спорообразовании раздуваются. Увел. Х2700.
Неспоровые сульфатредуцирующие бактерии из рода Desulfovibrio четко отличаются от спорообразующих по проценту Г + Ц в ДНК, наличию пигмента десульфовиридина и цито-хрома, монотрихиальным жгутикованием и отсутствием спорообразования.
Род Desulfotomaculum включает четыре вида. D. nigrifleans — типовой вид. Споры овоидные, субтерминальные. Бактерии могут расти без сульфатов на среде с пируватом. Глюкозу и другие углеводы не используют, термофильные. Температурный оптимум 55° С. Процент Г + Ц в ДНК 49,2. D. ruminis. Споры овоидные, терминальные или субтерминальные. Бактерии способны расти без сульфатов на среде с пируватом. Глюкозу и другие углеводы не сбраживают, мезофилы, Г + Ц в ДНК-49,2%.
D. orientis. Споры круглые, могут располагаться в клетках центрально, эксцентрально и терминально. Глюкозу и другие углеводы бактерии не сбраживают, не растут на средах с пируватом без сульфатов, мезофилы. Процент Г+Ц в ДНК 45,1. Японские авторы описали еще один вид спорообразующих сульфатредуцирующих бактерий — D. antarcticum. Споры этих бактерий овоидные, располагаются в клетках центрально или терминально. Нуклеотидный состав ДНК неизвестен. Этот вид отличается от других видов рода Desulfotomaculum способностью использовать глюкозу и разжижать желатин.
Аэробные спрообразующие бактерии. Род бациллюс (BACILLUS)
Аэробные спорообразующие бактерии составляют довольно обширную группу микроорганизмов. Они широко распространены в природе и играют большую роль в разнообразных биологических процессах. С использованием этих бактерий в промышленности освоено производство ценных ферментов, антибиотиков, органических кислот и других соединений.
Среди этой группы бактерий имеются и патогенные для человека и животных формы. В этом отношении особенно тщательно изучен сибиреязвенный бацилл. Имеются данные о фитопатогенных свойствах отдельных видов спорообразующих бактерий, обусловливающих развитие гнили и других поражений у растений.
Некоторые виды спорообразующих бактерий являются возбудителями болезней полезных насекомых. Образуя единственные в своем роде покоящиеся формы организмов — споры, эти бактерии служат объектом многочисленных исследований для выяснения природы ана- и парабио-тического состояния живой материи. С другой стороны, благодаря образованию спор, обладающих среди известных форм живых организмов наиболее высокой устойчивостью к неблагоприятным воздействиям внешней среды, спорообразующие бактерии представляют собой основной фактор биологического разрушения пищевых продуктов и различных других материалов.
Борьба со спорообразующими бактериями является одной из главных практических задач в консервной промышленности, при консервации крови и многих других процессах хранения и переработки сельскохозяйственных и пищевых продуктов.
Спорообразующие бактерии распространены повсеместно — в воздухе, водоемах, на растительных и животных остатках и других естественных субстратах.
Благодаря способности образовывать споры, обладающие большой устойчивостью к внешним воздействиям, спорообразующие бактерии остаются жизнеспособными при самых неблагоприятных условиях. Эти формы микроорганизмов выдерживают резкие колебания температуры, отсутствие влаги и воздуха, действие различных химических соединений, обычно губительно влияющих на живые организмы. Поэтому среди бесчисленного множества самых различных форм микроорганизмов спорообразующие бактерии являются наиболее устойчивыми и приспособленными к изменениям среды. Часто при анализе разных субстратов, подвергшихся неблагоприятным воздействиям, единственными представителями микробов бывают зародыши спороносных бактерий — споры.
Таким образом, спороносные бактерии имеют по сравнению с другими формами микробов большие возможности приспосабливаться к тем или другим условиям среды.
Спорообразующие бактерии распространены в морях и океанах, включая их глубокие слои и арктические воды. Работами А. Е. К рисса, С. И. Кузнецова и других советских ученых установлено распространение разнообразных видов спорообразующих бактерий в водоемах и их участие во многих биологических процессах среды своего обитания.
Большинство видов спорообразующих бактерий, подобно другим микроорганизмам, в основном обитает в почве. Однако не все почвы богаты спороносными бактериями. Содержание их в одной и той же почве различно в зависимости от окультуренности почвы, метеорологических, сезонных изменений и многих других факторов.
Почвы разных типов или возраста содержат неодинаковое число спорообразующих бактерий. Значительная часть таких бактерий обитает в бурых, сероземных, пустынных почвах, характеризующихся малым содержанием органического вещества. В богатых перегноем почвах спорообразующих бактерий сравнительно мало, например в арктических, дерново-подзолистых, лесных и горнолуговых.
Наряду с изменениями в общем (валовом) содержании спорообразующих бактерий особенно большие различия отмечаются в их видовом составе. В одних типах почв преобладают одни виды, в других — другие.
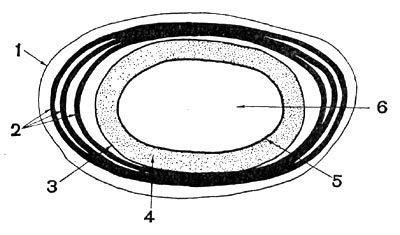
Рис. 65. Схематическое изображение строения споры: 1 — экзоспориум; 2 — слои споровой оболочки; 3 — внешняя мембрана споры; 4 — кора; 5 — внутренняя мембрана споры; 6 — сердцевина.
Фундаментальными работами в области экологии аэробных спорообразующих бактерий явились исследования Е. Н. Мишустина и его сотрудников. Были выявлены определенные закономерности распространения в почве отдельных видов спороносных бактерий. Так, установлено, что в биологических процессах, протекающих в почве, неспороносные бактерии связаны с превращениями гумуса на первых стадиях его минерализации, и поэтому по соотношению числа спороносных и неспороносных бактерий можно судить об интенсивности минерализации органического вещества в почве. Были выявлены характерные группировки спорообразующих бактерий, свойственные определенным почвенным типам и эколого-геогра-фическим зонам. На основе этих работ развились представления о показательных свойствах спорообразующих бактерий, отражающих почвообразовательный процесс, и описаны доминантные и типичные группы этих организмов для отдельных почвенно-климатических зон.
Закономерности эколого-географического распространения в почве различных видов споро-образующих бактерий достаточно рельефно обнаруживаются и по вертикальной зональности — при обследовании горных районов с большим разнообразием типов почвы, растительных формаций и климатических условий. С увеличением высоты в горах содержание и состав бактерий претерпевают закономерные изменения, в общем подобные тем, которые отмечаются при обследовании разных типов почв по широтной зональности.
Таким образом, изучение эколого-географического распространения спорообразующих бактерий выявляет специфику микробного населения различных типов почв и подтверждает учение В. В. Докучаева о зонах природы, горизонтальной и вертикальной зональности почвенного покрова. Общее количество и видовой состав споро-образующих бактерий в почве подвержены большим изменениям в зависимости от ее влажности и температуры.
При повышении влажности почвы после обильного дождя отмечается интенсивное размножение неспороносных бактерий. Их массовое развитие приводит к угнетению роста и развития спороносных бактерий. Их количество в почве может уменьшиться. Такое явление наблюдается при влажности почвы в пределах 50—60% и более от полной влаго-емкости. При умеренной влажности, в особенности в несколько подсохшей почве, количество спороносных бактерий увеличивается, а число неспороносных бактерий значительно уменьшается.
Много разнообразных спорообразующих бактерий обнаруживается в почвах со сравнительно высоким температурным режимом. Обильны бактериями почвы южных районов, а на севере и в почвах полярного круга их сравнительно мало. Значительный перегрев почвы на юге губительно действует на неспороносные бактерии, в то время как споровые формы переносят его хорошо.
У хорошо известного вида спорообразующих бактерий — Вас. mycoides установлено наличие эколого-географических разновидностей, характерных для отдельных почвенно-климатиче-ских зон. Выявлено, что у культур этих бактерий с удалением от севера к югу положение оптимальной и максимальной температур развития соответственно повышается. Южные культуры бактерий интенсивнее размножаются и обладают гораздо более высоким осмотическим давлением в клетках, нежели северные. У других видов спороносных бактерий такой приспособительной реакции к температурным условиям среды не отмечено.
Своеобразие эколого-географических условий приводит к расселению отдельных видов и разновидностей спорообразующих бактерий в определенных типах почв. Так, установлено очаговое распространение в красноземах неестественных, правозавитых, или так называемых инверсивных, форм Вас. mycoides. В солончаках обнаружены особые, галофильные разновидности спорообразующих бактерий, приспособившиеся к жизни в среде с высокой концентрацией солей. На питательных средах с небольшими концентрациями минеральных солей эти культуры развиваются очень слабо. Растительный покров оказывает значительное влияние на количественный и качественный состав спорообразующих бактерий в почве.
В почве, непосредственно прилегающей к корням растений (ризосфера), основная масса микробов представлена неспороносными бактериями. Споровых форм здесь очень мало. Однако на разлагающихся растительных остатках, которых много в почве осенью, спорооб-разующие бактерии обнаруживаются в значительном количестве.
Многие виды спорообразующих бактерий активно продуцируют ферменты, разлагающие обычно трудно разрушающиеся растительные остатки. Благодаря деятельности микроорганизмов почва обогащается ценными питательными веществами и освобождается от многих вредных организмов и продуктов. Возделывание в течение ряда лет одной и той же культуры может значительно изменить качественный состав спорообразующих бактерий в почве. Сдвиги в видовом составе споровых бактерий отмечаются при многолетнем культивировании люцерны и хлопчатника. В севооборотах эти различия несколько стираются, однако для микробиологических исследований как споровых, так и других форм бактерий в разных почвах знание истории поля обязательно. Накопление в почве спорообразующих бактерий-антагонистов оказывает благотворное действие на ее оздоровление, приводя к снижению в почве количества болезнетворных микробов.
Установлена ведущая роль спороносных бактерий в более глубоких стадиях распада веществ в компостах и других органических, соединений почвы. Обработка почв и агрохимические мероприятия, приводящие к уменьшению содержания подвижного органического вещества в почве, сопровождаются повышением содержания бациллярной флоры. Подобное явление отмечается и при длительном использовании минеральных удобрений. Количество спорообразующих бактерий повышается в нижних слоях почвы.
Сравнительно высокое содержание в почве спор бактерий является отражением определенной стадии в развитии этих организмов и не может свидетельствовать о неактивности спороносных бактерий.
О спорах бактерий, их химическом составе и структуре, процессах образования и прорастания подробно было рассказано в предыдущей главе. Здесь же мы остановимся на отличительных чертах, характерных для аэробных спорообразующих бактерий.
Поверхность наружной оболочки спор разнообразна у аэробных бактерий: либо гладкая, либо с выростами, выступами и почками. Оболочка составляет значительную часть споры. Основными ее компонентами являются белки (60—90%) и липиды. Состав аминокислот в белках оболочки неодинаков у бактериальных спор разных видов, однако большая часть цистина, содержащегося в значительном количестве в спорах, обнаруживается именно в оболочке.
Под оболочкой располагается кора споры (рис. 65), состоящая в основном из муреина. который является нерастворимым структурным материалом коры.
В течение многих лет покоящаяся спора рассматривалась как почти инертное тело, обладающее несколькими ферментами или вовсе лишенное их. Некоторых ферментных систем, имеющихся в вегетативной клетке, например цитохромов и ферментов цикла трикарбоновых кислот, ничтожно мало в споре. Это указывает на то, что в спорах аэробный энергетический метаболизм осуществляется иным путем, чем в клетках. Аналогичные ферменты спор и вегетативных клеток в спорах более термоустойчивы.
Споры аэробных бактерий обладают различной термоустойчивостью, что обусловлено как видовыми особенностями, так и условиями их образования. Установлено, что споры, образующиеся при высокой температуре, а также полученные на средах с высоким содержанием кальция, парааминобензойной кислоты и некоторых жирных кислот, более жароустойчивы.
Споры более устойчивы, чем вегетативные клетки, от которых они образовались, к воздействию проникающей радиации, ультразвука, гидростатического давления, замораживания, разрежения и др. Устойчивость спор к кислотам и щелочам во многом зависит от их принадлежности к различным видам бактерий. Споры некоторых культур бактерий остаются жизнеспособными даже при кипячении в концентрированной соляной кислоте в течение 20 мин. При этом резистентность спор во много раз повышается при их обезвоживании.
К спиртам и другим органическим растворителям устойчивость спор намного выше, чем у вегетативных клеток. Споры менее устойчивы к действию алкилирующих агентов. При воздействии комплекса определенных ферментов удается достигнуть растворения спор.
Механизм устойчивости спор к действию высокой температуры, проникающей радиации и других физико-химических факторов полностью еще не выяснен. Термоустойчивость спор объясняется наличием кальция, дипиколиновой кислоты и в некоторой степени диаминопимелиновой кислоты. Эти вещества определяют в значительной мере и резистентность спор к действию радиации. Полагают, что споры содержат комплекс особых веществ, подобных хелатным соединениям, которые предохраняют жизненно важные структурные элементы от губительного воздействия физико-химических факторов. Важным фактором, приводящим к образованию спор, является отсутствие для данного вида аэробных бактерий благоприятных условий для роста и развития. Способствует спорообразованию обеднение питательной среды.
У аэробных бактерий анаэробные условия выращивания подавляют как спорообразование, так и рост. Спорообразование аэробных бактерий протекает лишь в условиях доступа кислорода. Интенсивность споруляции подавляется в разной степени при выращивании бактерий в неблагоприятных условиях — при температуре выше оптимальной или реакции среды менее подходящей для обильного развития. Неблагоприятные условия роста, ускоряющие споруляцию бактерий, как правило, не обусловливают обильного «урожая» спор.
Разные виды аэробных бактерий отличаются расположением и формой спор. Эти признаки имеют важное систематическое значение. У некоторых видов расположение спор строго определенное — центральное, околоконцевое и концевое. У других видов не наблюдается строгой локализации. Высокое содержание в питательной среде источников азотного и углеродного питания, обеспечивающих максимальный рост бактерий, может подавлять индукцию спорообразования. Так, потребность в глюкозе — основном энергетическом источнике вегетативного роста аэробных бактерий — является несущественной для спорообразования некоторых видов, а в отдельных случаях она угнетает процесс споруляции. Культуры определенных видов бактерий более интенсивно спорулируют при уменьшении в среде аланина, валина, лейцина и изо-лейцина. У других бактерий отсутствие в среде лейцина и серусодержащих аминокислот угнетает спорообразование.
Интенсивность процесса образования спор в значительной мере обусловлена наличием в среде минеральных солей, содержащих ионы кальция, магния, калия, марганца, железа, цинка, меди и некоторых редких металлов (кобальта, лития, кадмия, никеля). Оптимальная концентрация минеральных солей и металлов различна в зависимости от вида выращиваемой культуры бактерий, условий ее роста. Роль минеральных солей и отдельных ионов металлов настолько велика, что в их отсутствии спороношение может вовсе не наблюдаться. Изменения концентраций их в питательной среде могут оказать иногда резко различное действие.
Спорообразование отдельных культур бактерий настолько зависит от наличия минимальных концентраций некоторых металлов, что эта особенность используется в аналитических целях.
Роль минеральных солей различных металлов в образовании бактериальных спор установлена недавно, и многие вопросы механизма их действия пока не выяснены. Полагают, что их специфическое действие связано с активированием различных ферментативных систем. Значение таких элементов, как кальций, заключается также и в том, что они способствуют образованию разнообразных низкомолекулярных веществ в спорах, обеспечивающих их термостабильность.
Сравнительно слабо изучено влияние на образование спор различных витаминов и других ростовых веществ. Некоторое усиление этих процессов отмечено под влиянием параамино-бензойной и фолиевой кислот.
В результате всестороннего изучения биохимии спорообразования показано, что при спору-ляции не образуются новые ферментные системы. Аналогичные ферменты, обнаруживаемые в вегетативной клетке и споре, имеют одинаковые молекулярный вес, аминокислотный состав и другие физико-химические свойства. Теплоустойчивость ферментов спор вызвана присутствием в них ионов кальция, марганца и других металлов. Данные исследований свидетельствуют о том, что природа и функция ферментативного аппарата, осуществляющего синтез белков в споре и материнской вегетативной клетке, одинаковы. Основной системой транспорта электронов в спорах является растворимая флавопротеино-ксидаза. При споруляции выявлена высокая активность ряда ферментов, например аргиназы, некоторых нуклеаз. Основная масса белков споры синтезируется заново в период споруляции. Одновременно отмечается процесс распада белков, причем многими авторами обнаружена тесная взаимосвязь протеолитической активности с функцией спорообразования у бактерий. Из вновь синтезированных белков лишь незначительная часть используется для создания структурных элементов споры. В целом споруляцию можно охарактеризовать как процесс интенсивного распада белков и всех основных типов РНК материнской клетки с одновременным образованием новых белковых компонентов.
Различия в составе белков споры и вегетативной клетки выявляются серологическими методами. По антигенной структуре спорулирующая и аспорогенная формы культур одного и того же вида различны. Не установлено существование какого-либо антигена, общего для спор бактерий разных видов. Наоборот, по антигенной структуре имеется достаточно отчетливая дифференциация между спорами разных видов.
В способе прорастания споры и продолжительности процесса также отмечается определенное постоянство, характерное для различных видов бактерий. Старые споры прорастают в течение более длительного периода, чем молодые. Время прорастания молодых спор является довольно постоянным у различных бактерий и может служить видовым признаком. Если перенести только что сформировавшиеся споры в благоприятную питательную среду, то прорастание их начинается не сразу, а по истечении 2—3 ч.
Установлено, что на начальной стадии прорастания спор происходит выделение в среду так называемого экссудата прорастания, характеризующегося специфическим поглощением ультрафиолетовых лучей. Вещество это было выделено и идентифицировано как кальциевая соль дипиколиновой кислоты (пиридин — 2,6-дикарбоксиловая кислота). Дипиколинат кальция составляет приблизительно половину сухой массы экссудата прорастания, что соответствует 15% от общей сухой массы покоящихся спор. Остальная часть экссудата прорастания представлена свободными аминокислотами, высокомолекулярными и низкомолекулярными белковыми веществами. Обнаружено особенно большое количество недиализируемого гексозаминсодержащего пептида с молекулярным весом порядка 10 000, в состав которого входят диаминопимелиновая кислота, аланин, глюта-миновая кислота и гексозамины. Условно это соединение названо ДАП-гексозаминпептид. Это соединение входит в состав оболочки спор, в отличие от дипиколиновой кислоты, обнаруживаемой внутри спор.
Среди факторов, способствующих процессу прорастания спор, большое внимание уделяется тепловой активации, действие которой более выражено при наличии в среде некоторых химических соединений. Тепловая активация достигается предварительным кипячением суспензии спор при 97—100° С в течение 10—15 мин или при 65° С в течение 2 ч. Иногда этот эффект достигается при обычной пастеризации. Тепловая активация прорастания спор носит общий характер для разных видов спороносных бактерий, хотя она более специфична для аэробов, чем анаэробных видов. Обычно тепловое воздействие усиливает и ускоряет прорастание спор в течение короткого промежутка времени. Действие подобной обработки спор, как правило, преходяще, хотя при этом отмечается и некоторое изменение условий, необходимых для течения процесса прорастания спор. Так, для лучшего прорастания спор сенного бацилла без предварительной тепловой обработки необходимо присутствие в среде L-аланина, аденозина, глюкозы и тирозина, тогда как после тепловой обработки оптимальные условия прорастания спор обеспечивают наличие лишь глюкозы и аденозина. Установлено также, что сохранение в течение длительного времени спор при 37° С после их тепловой активации ведет к деактивации. Тепловая обработка этих спор после длительного хранения вновь ведет к реактивации прорастания.
Имеются данные о некотором активирующем влиянии на прорастание спор умеренного воздействия ультразвука. Исследования последних лет показали, что интенсивность прорастания спор специфически повышается в присутствии некоторых веществ, в первую очередь L-аланина, пуриновых нуклеозидов и глюкозы. Иногда необходимо наличие, помимо L-аланина, ряда других аминокислот. Из рибозидов наиболее эффективным оказался аденозин.
Алании является наиболее специфическим активатором процесса прорастания спор из всех известных соединений, обладающих подобным действием. Оптимальные условия для прорастания спор анаэробных бактерий несколько более сложны, хотя факторы, активирующие процессы прорастания спор аэробных видов, и в этом случае оказывают активирующее действие.
Наряду с активаторами выявлен ряд веществ, угнетающих процесс прорастания спор. Прорастание спор у аэробных видов угнетается в присутствии оксина, некоторых мышьяковистых соединений, углекислого натрия, высоких концентраций ряда аминокислот — глицина, метионина, цистеина и валина. Характерно угнетающее действие D-аланина в отличие от L-аланина.
Механизм активирующего и угнетающего действия ряда факторов на прорастание спор остается неясным. Одни авторы приписывают активаторам определенную роль в процессах деполимеризации различных составных элементов в спорах, высвобождении и активации ряда ферментных систем, обусловливающих начало прорастания спор. Другие считают их энергетически активными соединениями, стимулирующими биохимические процессы в момент прорастания спор.
Обычно не все споры, вносимые в благоприятную среду, прорастают сразу. Некоторое их количество остается непроросшими в течение различного времени, не теряя, однако, своей жизнеспособности. Установлено, что при длительном хранении процент прорастающих спор постепенно снижается. Причина этого явления и природа подобных «дремлющих» спор остаются неясными.
В покоящейся споре имеются ферментные системы, выявляемые лишь после их активации под влиянием определенных факторов и химических соединений, которые, однако, исключают прорастание спор. К таким системам относятся прежде всего окислительные ферменты.
Активация их достигается обработкой спор незначительными количествами L-аланина, аденозина и некоторыми другими веществами, а также кипячением или пастеризацией суспензий спор. Перевод неактивной ферментной системы в активную без признаков прорастания спор считается первым этапом биохимических изменений в спорах, ведущих в дальнейшем к началу процесса их прорастания. Полагают, что под действием активаторов дипиколиновая кислота, связывающая окислительные и другие ферменты, подвергается деполимеризации с освобождением ферментных систем.
С помощью высокочувствительного микрореспирометра у бактериальных спор установлена дыхательная активность.
При изучении химического состава клеточной стенки спор различных видов аэробных бактерий обнаружены характерные различия в содержании ДАП-гексозаминпептида и других веществ — полисахаридов, липидов. ДАП-гек-созаминпептид высвобождается из стенки спор под действием литического агента, уподобляемого по своим свойствам лизоциму.
Классификация аэробных спорообразующих бактерий
По современным представлениям, аэробные спорообразующие бактерии, или бациллы, объединяются в отдельный род Bacillus семейства Bacillaceae. Этот род, включающий много разнообразных видов, имеет ряд характерных особенностей и отличается от других бактериальных организмов комплексом морфолого-физио-логических признаков, из которых наиболее важными являются палочковидная форма клеток, способность образовывать эндоспоры, потребность в свободном кислороде для роста.
Общая краткая характеристика объединяемых в род бацилл аэробных спорообразующих бактерий сводится к следующему.
Бациллы — свободноживущие, одноклеточные, нефотосинтезирующие, аэробные, палочковидные клетки, образующие типичные эндоспоры. Относятся к гетеротрофным организмам. Размножаются поперечным делением клеток. Ветвление и почкование клеток как способ размножения не отмечены. Поперечный размер клеток варьирует в пределах 0,4—2 мкм. Вегетативные клетки имеют вид прямых или слабоизогнутых палочек с параллельными сторонами и округлыми концами, которые в редких случаях резко обрезанные.
Расположение клеток различное — от одиночных до длинных цепочек. За исключением сибиреязвенного бацилла, бактерии этого рода подвижные, снабжены перитрихиально расположенными жгутиками.
Большинство видов является грамположительными бактериями, часть — грамвариабильными. Клетки хорошо окрашиваются обычными анилиновыми красками. Ни один представитель рода не является типично кислотоустойчивым организмом. У многих видов отмечается наличие внутриклеточного жира, гликогена, волю-тина и других включений. Капсула встречается лишь у сибиреязвенного бацилла и некоторых других видов при специфических условиях роста.
В бактериальных клетках в общем количестве оснований ДНК 32—65 мол.% гуанина и цитозина.
Большинство видов, исключая некоторые, главным образом энтомопатогенные, формы, хорошо растет на мясопептонном агаре (МПА) при реакции среды, близкой к нейтральной. Отдельные виды развиваются в щелочной среде и требуют особых источников азота или углерода.
Культуральные особенности видов, выросших на разных средах, резко различны. На твердых питательных средах образуются колонии от 1—2 до 5 мм и более в диаметре: гладкие, зернистые, пленчатые, складчато-морщинистые и сухие, слизеобразующие и пастообразные с характерной структурой края. При развитии на жидких средах обнаруживается тенденция к образованию поверхностной пленки. Встречаются виды, образующие на поверхности агаризованных сред подвижные колонии.
Большинство видов бактерий активно продуцируют ферменты, гидролизующие белки, крахмал и другие субстраты. Многие виды обладают антагонистическими свойствами и вырабатывают антибиотики полипептидной природы. Отдельные виды нуждаются в присутствии витаминов, аминокислот и других дополнительных факторов роста. Кислоту и газ продуцируют лишь бактерии некоторых видов, все остальные при росте на углеводах образуют одну кислоту. Оптимальная температура роста обычно варьирует в пределах 30—40° С. Встречаются виды, развивающиеся при температуре ниже 12° С и выше 50° С.
Некоторые аэробные бактерии — возбудители болезней. Вас. anthracis вызывает сибирскую язву у человека и животных; Вас. larvae — возбудитель американского гнильца медоносной пчелы; Вас. alvei и Вас. pulvifaciens трактуются как организмы, играющие определенную роль в болезнях пчел; Вас. popilliae и Вас. lentimorbus — возбудители молочной болезни японского жука; некоторые виды группы Вас. cereus-thuringiensis вырабатывают специфические энтомоцидные токсины.
Классификация видов аэробных спорообразующих бактерий разработана недостаточно полно. Одной из причин этого является ограниченность различий во внешних признаках бактерий. Известно, что большинство видов различаются между собой малозначимыми признаками строения и развития клеток, по форме колоний, а также физиологическим признакам. Многие вопросы биологии спороносных бактерий требуют глубоких исследований. Хорошо изучены только те виды, которые имеют практическое значение (патогенные, бродильные формы, продуценты различных антибиотиков, ферментов и т. д.). Вопросы систематики спорообразующих бактерий изучались специалистами прикладных наук для диагностических целей, в основном санитарно-гигиенического назначения.
Для выделения бактерий в самостоятельные виды одни авторы считают решающими ферментативные свойства организмов, другие — морфологические и культуральные признаки, третьи — цитологические особенности бактериальной клетки и т. д. Во всех подобных случаях принципы систематики меняются по мере выявления новых особенностей в строении и физиологических свойствах организмов и использования их в качестве признаков видовой идентификации. Систематика разных видов спорообразующих бактерий отражает степень их изученности, а следовательно, должна развиваться и изменяться по мере накопления новых данных.
Вопросы систематики бактерий представляют исключительную важность для работ по изысканию и изучению образования разнообразных физиологически активных веществ микробного происхождения. Биологический вид представляет единство специфических морфологических и физиолого-биохимических признаков организма, определяющих все особенности его жизнедеятельности, распространения и взаимодействия с внешней средой. В связи с этим образование того или иного продукта жизнедеятельности, как и характерные биологические особенности, невозможно представить в отрыве от видовой принадлежности микроорганизма.
Работы в области систематики спорообразующих бактерий и определения их положения в мире микроорганизмов начались еще до открытия спор у бактерий. Термин Bacillus на разных этапах развития микробиологии использовался вольно для обозначения как спороносных, так и неспороносных бактерий. В сущности, подобное положение сохранилось до наших дней, и мы нередко затрудняемся не только идентифицировать описанные виды, но и установить их истинную принадлежность к спорообразующим бактериям. Многие спорообразующие бактерии именовались различными авторами под родовыми названиями, которые в настоящее время служат обозначением родов неспороносных бактерий и других микроорганизмов, что еще более осложняет точное представление об описываемом организме.
Таксономия бактерий раннего периода в основном была конструктивной и имела диагностическое назначение. На этом этапе рассматривались подходы к созданию крупных систематических категорий.
По мере накопления фактического материала начали предпринимать попытки классификации по аналитическому принципу, учитывая родственные взаимоотношения микроорганизмов.
Систематика спорообразующих бактерий развивалась по мере эволюции взглядов на принципы дифференциации и идентификации микробных видов, а также усовершенствования методов морфолого-физиологического изучения бактерий.
Наиболее ранними в этом отношении были попытки микробиологов разграничить виды спорообразующих бактерий по морфологическим признакам, в частности по форме спор, способу их прорастания и т. п. Малозначимость этих признаков дифференциации и их неспецифичность привели к тому, что в дополнение к предыдущим описаниям было идентифицировано много новых видов, большинство из которых в последующем не были приняты или трактуются сейчас как синонимы других систематических категорий.
Широкое распространение спорообразующих бактерий в окружающей среде и их частое обнаружение в различных продуктах, особенно при порче, вызвали к ним определенный интерес. Первые работы по классификации этой группы бактерий были выполнены в основном представителями пищевой и санитарной микробиологии. При этом одни авторы основывались на морфологических признаках микроорганизмов, а другие — на физиологических. Классификация, основанная на различиях в способности аэробных спорообразующих бактерий сбраживать субстраты, предусматривала подразделение этих организмов на три группы по способности сбраживать глюкозу и образовывать ацетилметилкарбинол. Согласно другой классификации, которую в настоящее время используют наиболее часто, данный род бактерий разделяется на три группы по соотношению поперечных размеров спор и вегетативных клеток. Группа I — наиболее обширная — включает виды спороносных бактерий, у которых не отмечается отчетливого раздувания спорангия в процессе образования спор. Группа II объединяет бактерии, образующие овальные споры, раздувающие спорангий. Группа III охватывает бациллярные виды, характеризующиеся округлыми или шаровидными спорами, раздувающими спорангий; объединяет редко встречающиеся виды спорообразующих бактерий.
Вопросы классификации различных видов аэробных спорообразующих бактерий разрабатывались многими отечественными авторами. Особенно тщательно были изучены морфолого-физиологические особенности отдельных групп и видов этих бактерий Е. Н. Мишустиным с сотрудниками. Данные этих исследований свидетельствуют о большом многообразии видов и экологических разновидностей спорообразующих бактерий в зависимости от мест их обитания, почвенно-климатической зональности и микробного ценоза разных типов почв.
В настоящее время для определения видов спорообразующих бактерий наряду с морфоло-го-физиологическими особенностями используют и многие другие признаки.
Важными критериями для определения и дифференциации бактерий являются отношение к действию специфичных фагов, агглютинация с гомологичными сыворотками к споровому, соматическому и жгутиковому антигенам, рост при высокой концентрации солей, различной температуре и т. п. Многие отмеченные особенности применяются для внутривидового подразделения и выделения культур спороносных бактерий в отдельные разновидности, серотипы и другие мелкие систематические подразделения. В ряде случаев некоторые признаки служат основой для выделения культур спорообразующих бактерий в новые виды. Так, например, в качестве новых видов описаны психрофильные, развивающиеся при низкой температуре культуры спорообразующих бактерий. Подобным же образом выделены некоторые активные продуценты амилолитических и целлюлолитических ферментов. В литературе было описано много новых видов бацилл, патогенных для некоторых насекомых, растений и животных, что в большинстве случаев не подтвердилось в последующие годы.
Успехи в области создания счетно-вычислительной техники дали основу для быстрой математической обработки данных по родству и отдаленности организмов. Применение этой техники в бактериальной таксономии получило название числовой таксономии и было введено в микробиологию в последние годы. Числовая таксономия рассматривает группирование и идентификацию организмов с учетом их подобия при анализе не менее чем 50—60 объективных признаков. Этот метод с разработкой схем подобия и разграничения уже использован рядом авторов для систематики различных видов спороносных бактерий.
Особого внимания заслуживает разработка новых подходов к трактовке вопросов филогенетической таксономии бактерий с помощью биохимических исследований. Наибольший интерес в этом отношении представляют попытки классификации бактерий с использованием новейших данных молекулярной биологии.
Биология аэробных спорообразующих бактерий
Клетки аэробных спорообразующих бактерий имеют палочковидную форму и в сравнении с неспороносными бактериями, как правило, более крупных размеров. Вегетативные формы спороносных бактерий обладают более слабым активным движением, хотя им присущи пери-трихиально расположенные жгутики. Длина клеток вегетативных форм варьирует, особенно в зависимости от возраста культуры. У некоторых видов спороносных бактерий в старых культурах отмечается образование характерных полиморфных форм. Размеры и форма бактериальных клеток подвергаются значительным изменениям в процессе образования спор, что во многих случаях может служить дополнительным признаком для распознавания отдельных видов. В молодых культурах содержимое клеток однородно, по мере старения гомогенность исчезает, появляется зернистость. При этом отмечается значительное накопление внутриклеточного жира, который выявляется при специфическом цитохимическом окрашивании в виде зерен различной формы. Количество и характер свободного внутриклеточного жира различны у разных видов спороносных бактерий и, по мнению ряда авторов, могут быть использованы для определения видовой и групповой принадлежности бактерий.
Форма и величина спор — постоянные признаки — имеют первостепенное значение при групповом и видовом разграничении спорообразующих бактерий. Немаловажное значение при этом придается соотношению поперечных размеров спор и клеток, а также локализации спор. Спорулирующая клетка, или спорангий, у отдельных видов бактерий раздувается в месте расположения спор, если поперечник спорангия больше поперечника клетки. В зависимости от расположения спор, раздувающих спорангий, клетки принимают либо плектридиальную, либо клостридиальную форму.
Споры бактерий в благоприятных условиях прорастают, причем в характере прорастания отмечается постоянство, присущее определенным видам. Различают полярное и экваториальное прорастание спор. Как редкое исключение наблюдается косое прорастание. Сказанное относится к спорам продолговатой формы. Наблюдения показали, что некоторые виды бактерий имеют строго определенный тип прорастания спор; у отдельных видов наблюдаются и отклонения. Помимо места, откуда происходит прорастание, для различных видов характерными являются отдельные цитологические изменения, происходящие в процессе прорастания и сбрасывания (разрывания или пробивания) оболочки споры. Эти особенности прорастания спор впервые были отмечены в конце прошлого столетия И. И. Мечниковым.
Для различных видов спороносных бактерий характерны культуральные особенности роста на питательных средах. При росте на твердых средах на поверхности образуются своеобразные скопления клеток в виде отдельных бактериальных колоний. Строение колоний зависит от состава питательной среды и условий культивирования, особенностей биохимической деятельности и химического состава бактерий, интенсивности их роста, особенностей движения после деления.
Форма поверхности колоний спорообразующих бактерий самая разнообразная: гладкая жирноблестящая, плоская зернистая или крупинчатая, пленчатая влажноблестящая, сухая мучнистая, складчато-морщинистая и т. п. Один и тот же вид спороносных бактерий образует на средах различного состава разнообразные колонии. Поэтому при описании культуральных особенностей выращиваемых бактерий строго придерживаются определенного состава питательных сред и условий выращивания.
Для большинства спорообразующих бактерий очень подходящи среды, содержащие усвояемые углеводные и сложные органические соединения. С этой целью успешно могут быть использованы мясо-пептонный агар с глюкозой, сахарозой (1—2%), сусло-агар и т. п. Некоторые виды спорообразующих бактерий образуют на твердых питательных средах подвижные колонии. Природа этого явления пока недостаточно изучена, но показано, что оно связано с усиленной подвижностью клеток и некоторыми физико-химическими особенностями питательной среды.
Важным диагностическим признаком для определения видов спороносных бактерий является строение края колоний. Одни виды образуют колонии с ризоидными, бахромчатыми и ветвистыми краями, другие — с гладкими округлыми или бугристыми, грубо волнистыми. Подобные особенности в ряде случаев удается подметить невооруженным глазом, но особенно хорошо они выявляются при микроскопии с небольшим увеличением.
Консистенция бактериальных колоний весьма разнообразна. Культуры группы картофельного и сенного бацилла характеризуются образованием плотных, вросших в агар, сухих морщинистых колоний. У других видов бактерий колонии пастообразные или слизеобразу-ющие, гладкие. Образование пигментов у спорообразующих бактерий отмечается сравнительно редко. Некоторые культуры бактерий при росте на белковых средах приобретают бурую пигментацию, диффундирующую в субстрат. Подобное окрашивание не связано с образованием пигмента, а вызвано продуктами распада сложных органических соединений.
При культивировании сибиреязвенного бацилла на жидких средах отмечается выделение пигментов красного и бурого цвета. У отдельных культур сенного бацилла и некоторых других видов наблюдается образование красного пигмента, который был идентифицирован как пульхеримин и пульхериминовая кислота.
При росте на жидких питательных средах одни виды спороносных бактерий образуют поверхностную пленку, другие — нет. Культуры Вас. mycoides образуют характерную паутиновидную пленку, которая в пробирке располагается в форме конуса основанием вверх. Культуры бактерий, характеризующиеся образованием поверхностной пленки, при потере спорообразования лишаются этой способности.
Культуральные особенности роста различных видов спорообразующих бактерий, равно как и других микроорганизмов, отражают их характерные физиолого-биохимические свойства, особенности деления, размножения и расположения клеток. Отличительные особенности распределения клеток в процессе их размножения в колониях бактерий на-иболее четко выявляются при наблюдении в сканирующем микроскопе (табл. 37). Аэробные спорообразующие бактерии относятся к гетеротрофам, т. е. к микроорганизмам, нуждающимся в готовых органических соединениях. Для огромного большинства споро-образующих бактерий лучшими источниками азотного питания являются белки и аминокислоты сложных органических соединений. На средах с минеральными соединениями азота большинство этих бактерий развивается слабо.
Спорообразующие бактерии обладают активными протеолитическими ферментами. Они разлагают сложные белковые соединения на аминокислоты, а затем и на более простые азотистые вещества и аммиак. Способность спорообразующих бактерий к энергичному разложению сложных органических соединений до простых продуктов распада стала основой для физиологической характеристики этой группы бактерий как активных аммонификаторов. Однако различные виды спорообразующих бактерий по-разному относятся к источникам азотистого питания. Так, например, культуры группы сенного и картофельного бацилла более энергично сбраживают углеводы с образованием разнообразных промежуточных соединений. Для наиболее распространенных видов спорообразующих бактерий лучшими источниками азотного питания оказались пептон, гидролизат казеина, автолизат дрожжей и мочевина.
Среди почвенных аэробных спорообразующих бактерий имеются виды, которые довольно хорошо развиваются на безазотистой среде. К ним относятся олигонитрофильные бактерии, обладающие способностью усваивать атмосферный азот. Эти микроорганизмы характеризуются слабой азотфиксирующей способностью, однако их накопление в почве может значительно содействовать обогащению ее азотом. В этом отношении всестороннее изучение данной группы микроорганизмов представляет большое значение для повышения плодородия почв.
Большинство видов спорообразующих бактерий, широко распространенных в почве, обладает способностью использовать в качестве источника азота нитраты. Благодаря этому они вызывают процессы восстановления нитратов (денитрификацию) и переводят их в состав органических азотистых соединений. Таким образом, спорообразующие бактерии усваивают азот из различных источников в неодинаковой степени. Одни предпочитают аммонийный азот, другие — азот аминокислот; многие хорошо потребляют азот нитратов, тогда как некоторыми он не усваивается или используется слабо.
Активная аммонифицирующая деятельность спорообразующих бактерий имеет важное значение для повышения плодородия почв. Некоторые авторы отметили заметное увеличение урожайности растений при внесении в почву активных аммонификаторов. Спорообразующим бактериям принадлежит важная роль в превращениях фосфора. Многими авторами было показано, что культуры различных видов спороносных бактерий активно переводят нерастворимые фосфорные соединения в растворимые. В Советском Союзе с применением культуры спорообразующих бактерий, активно минерализующей сложные органические соединения фосфора, было налажено промышленное производство бактериального удобрения — фосфобактерина. Этот препарат широко и успешно применялся для повышения урожайности многих сельскохозяйственных культур.
Установлено несколько механизмов разложения фосфорных соединений спороносными бактериями. В одном случае происходит растворение трикальцийфосфата, в другом — перевод фосфоритов в апатиты или фосфитов в фосфаты. Более распространенным является минерализация органофосфатов в результате ферментативной деятельности бактерий и выделения ими кислот. Результаты исследований показывают, что отмеченные свойства не приурочены к особым видам, обнаруживаются у культур многих видов и разновидностей спороносных бактерий, а также других микроорганизмов.
Отечественными авторами были выделены и изучены культуры спорообразующих бактерий, способные разрушать полевые шпаты, каолин и другие формы силикатов.
Среди спорообразующих бактерий обнаружены виды и разновидности, характеризующиеся выраженной специализацией биохимической активности. Так, культуры Вас. pasteurii обладают уреазной активностью и хорошо развиваются в сильно щелочной среде. Редкой формой бациллярных организмов является Вас. fastidiosus, который не способен усваивать сахара, аминокислоты, белковые и другие соединения. Бактерии этого вида потребляют лишь мочевую кислоту и аллантоин.
В качестве источника углеродного питания спорообразующие бактерии могут использовать как простые сахара, так и полисахариды. Из моносахаридов лучше всего они усваивают глюкозу, а из дисахаридов — сахарозу. Отношение разных видов спорообразующих бактерий к углеводам различное, что широко используется в исследованиях по систематике для определения видовой принадлежности микроорганизмов. При развитии на простых сахарах, в особенности на глюкозе, спорообразующие бактерии синтезируют ряд продуктов — молочную, уксусную и пропионовую кислоты, ацетилметилкарбинол и т. п. Газообразование отмечается только у культур нескольких видов. Многие виды спорообразующих бактерий сравнительно хорошо усваивают различные органические кислоты и спирты.
В группе аэробных спорообразующих бактерий немало видов и разновидностей, которые для своего роста и развития нуждаются в различных витаминах, аминокислотах и других ростовых факторах. Так, для культур сибиреязвенного бацилла наряду с аминокислотами необходим тиамин. Культуры облигатных энто-мопатогенных видов — возбудителей молочной болезни японского жука, американского гнильца медоносной пчелы и других болезней насекомых — нуждаются во многих аминокислотах и витаминах.
Культуры сапрофитных спорообразующих бактерий также в ряде случаев проявляют специфические потребности в витаминном и аминокислотном питании. Наиболее характерной для подобных организмов является зависимость их роста и развития от наличия в среде тиамина, биотина, фо-лиевой и никотиновой кислот. В качестве обособленного нового вида описаны культуры спорообразующих бактерий, нуждающиеся в пан-тотеновой кислоте. Термофильным формам спороносных бактерий присуща более выраженная зависимость роста и развития от аминокислот, витаминов и других дополнительных факторов роста.
Спорообразующие бактерии — активные продуценты многих ферментов.
Бактерии группы сенного и картофельного бацилла обладают выраженной протеолитиче-ской и амилолитической активностью. В настоящее время культуры этих видов спороносных бактерий широко используются во многих странах для промышленной выработки амилазы и различных протеаз. С давних времен в Японии большой популярностью пользуется национальное блюдо «натто», которое изготовляют из соевых бобов при их обработке культурой сенного бацилла.
Некоторые виды спорообразующих бактерий вырабатывают ферменты, разлагающие пектин и протопектин растений. Этим бактериям при-писывается определенная роль в создании комковатой структуры почв и в образовании клубеньков у бобовых растений.
Выраженные ферментативные свойства некоторых видов спорообразующих бактерий обусловливают развитие гнилей и побурение плодов, фруктов и других растительных продуктов. При благоприятных условиях размножения некоторых видов бацилл — продуцентов фосфолипаз и других ферментов — вызывается порча пищевых продуктов, которые могут служить причиной гастроэнтеритов и других пищевых отравлений.
Среди бактериальных организмов группа аэробных спорообразующих бактерий известна своими сильно выраженными антагонистическими свойствами в отношении большого числа микроорганизмов. Спороносные бактерии можно поставить в один ряд с лучистыми грибками, пенициллами и аспергиллами, представляющими собой основные источники получения антибиотических веществ.
Подавляющее большинство антибиотиков бактериального происхождения выделено из группы спорообразующих бактерий. Первый антибиотический препарат — грамицидин — выделен из культуры спороносной бактерии. Советскими учеными был получен антибиотик грамицидин С, который успешно используется при лечении многих инфекций. Из различных видов бацилл получены антибиотики альвеин, бациллин, бацитрацин, лихениформин, субтилин, циркулин и др. Особый интерес вызвало открытие полимиксинов — антибиотиков бациллярного происхождения, которые оказались высокоактивными против грамотрицательных бактерий. В ряде стран организовано промышленное производство отдельных препаратов антибиотиков: полимиксина В, колистина, бацитрацина.
Кроме терапевтического использования, бацитрацин применяется в качестве стимулятора роста животных.
В настоящее время из различных видов спорообразующих бактерий выделено более 60 антибиотиков, из которых некоторые были испытаны в клинической практике. Однако антибиотики, полученные из спорообразующих бактерий, изучены недостаточно.
За некоторыми исключениями, антибиотики из спорообразующих бактерий представляют собой вещества белковой природы — полипептиды. Они состоят из различных аминокислот, причем наиболее характерным является то, что в состав полипептидных антибиотиков входят неестественные аминокислоты с правой оптической конфигурацией. Некоторые антибиотики полипептидной природы имеют сравнительно более простой состав и несложную формулу, представляя весьма удобную модель для изучения строения белка. С другой стороны, полипептидные антибиотики привлекают большой интерес биохимиков, так как эти вещества могут быть использованы при изучении одной из центральных проблем современной биологии — выяснении причин, обусловливающих специфичность биологически активного белка.
По механизму действия продуцируемые спорообразующими бактериями антибиотики разделяются на три группы. Группа эдеинов, представляющих собой пептиды с сильно основными свойствами, специфически подавляет синтез ДНК. Группа бацитрацинов и родственных им антибиотиков представляет собой циклические пептиды, которые угнетают синтез оболочки клеток. В обширную группу бациллярных антибиотиков включают грамицидины, поли-миксины и родственные им соединения, которые специфически блокируют синтез структурных компонентов или функцию клеточной мембраны. Характерной особенностью этих антибиотиков является то, что они образуются после активного размножения бактерий в процессе спорообразования. Результаты генетических исследований свидетельствуют, что биосинтез бациллярных антибиотиков тесно связан с образованием протеазы, характерной для процессов спорообразования. Поэтому синтез антибиотиков отражает особенности биохимической дифференциации клеток спорообразующих бактерий.
Спорообразующие бактерии широко распространены в природе и играют большую роль в биологии почвы; изучение антагонистических свойств спороносных бактерий внесет много ценного в наши представления о роли антагонизма микробов в процессах самоочищения почвы, воды и других природных субстратов.
Энтомопатогенные аэробные спорообразующие бактерии
Спорообразующие бактерии, обладающие энтомопатогенным действием, относятся, как правило, к группе аэробных микроорганизмов. Имеются лишь единичные описания анаэробных энтомопатогенных спорообразующих бактерий. Энтомопатогенные аэробные Спорообразующие бактерии можно условно подразделить на две группы: облигатные формы и факультативные.
В группу облигатных входят виды бактерий— возбудителей болезней насекомых, специфически приспособившихся к условиям обитания в организме насекомых. Например, Вас. ро-pilliae и Вас. lentimorbus вызывают так называемую молочную болезнь японского жука; Вас. larvae — возбудитель американского гнильца медоносной пчелы.
Группа факультативных или условно энтомопатогенных спорообразующих бактерий весьма разнообразна по составу и включает большое число видов. Сюда относят ряд энтомо-фильных видов, часто встречающихся в микрофлоре насекомых и играющих важную роль в этиопатогенезе различных инфекций у насекомых, — Вас. alvei, Вас. orpheus, Вас. subrugosum и многие другие. Весьма условно могут трактоваться как факультативно энтомопатогенные микроорганизмы культуры широко распространенного вида Вас. cereus со многими разновидностями.
Особую группу составляют спорообразую-щие бактерии, объединяемые в вид Вас. thuringiensis, который включает многие разновидности спорообразующих бактерий, вырабатывающих особые энтомоцидные токсины кристалловидной природы. Эти бактерии используют для промышленного получения инсектицидных препаратов.
Наконец, как факультативно или, точнее, потенциально Энтомопатогенные формы должны рассматриваться многие виды спорообразующих бактерий, которые вызывают гибель насекомых при проникновении в организм и развитии в их гемолимфе. В основе энтомопа-тогенного действия спорообразующих бактерий, развивающихся в полости тела насекомых, лежит неспецифический механизм действия, связанный с продуцированием ими ряда ферментов. Энтомопатогенное действие этих бактерий может быть обусловлено или образованием ряда физиологически активных соединений, например токсичных антибиотиков, или специфическими метаболитическими взаимодействиями с организмом насекомых. Литература по энтомологии изобилует описаниями многих новых видов спорообразующих бактерий, трактуемых различными авторами как энтомопатогенные. В большинстве случаев при описании этих видов авторы руководствовались лабораторными наблюдениями над их патогенным действием на отдельных насекомых, а очень часто только тем, что эти микроорганизмы были выделены из погибших насекомых. Трактовка культур как новых видов сопровождалась поверхностным изучением их, а малозначимые отличительные признаки рассматривались как основа для выделения их из известных видов бактерий.
Однако анализ фактического материала, накопленного в этой области, показывает, что, за исключением упомянутых выше облигатно энтомопатогенных форм и некоторых других видов, большинство описанных энтомопатогенных спороносных бактерий является представителями ранее описанных видов и разновидностей.
Группа облигатных энтомопатогенных бактерий — Вас. popilliae-lentimorbus, Вас. larvae — характеризуется общими признаками, принципиально важными для установления их родства. Все они морфологически близки — при развитии вегетативных клеток образуются споры, раздувающие спорангий; грамвариабильны. Культуры этих видов требуют для своего развития присутствия в среде аминокислот, тиамина и биотина, очень трудно растут на обычных лабораторных средах. Это указывает на более узкую биохимическую специализацию этих видов.
Культуры энтомофильных видов — Вас. alvei, Вас. para-alvei, Вас. pulvifaciens — примыкают к указанным видам по общим признакам. Бактерии этих видов также являются витаминза-висимыми, хотя хорошо развиваются и на обычных белковых средах.
Вид Вас. orpheus (Вас. laterosporus) сравнительно мало изучен. Его систематическое разграничение основывается на своеобразном характере спорообразования, при котором зрелая спора практически представляет собой спорангий с остатком клетки, окаймляющей спору в виде полулуния. Подобное явление иногда обнаруживается и в культурах других видов.
Общепризнано биологическое и систематическое родство энтомопатогенных спорообразующих бактерий Вас. cereus, Вас. thuringiensis и многочисленных их разновидностей. Потеря способности образовывать энтомоцидные токсины лежит в основе отграничения Вас. thuringiensis от Вас. cereus. Бактерии этих видов имеют много общего по культуральным, морфологическим и физиологическим особенностям. Как правило, строго обязательной является потребность в присутствии аминокислот. Вита-минзависимые штаммы обнаруживаются у отдельных разновидностей. Паразитизм у энтомопатогенных бактерий сопровождается развитием у них биохимической дифференциации, выражающейся в более узкой специализации питания. Облигатные формы этих микроорганизмов характеризуются зависимостью от многих аминокислот, витаминов и других факторов роста, тогда как сапрофитные и условно патогенные энтомопатогенные виды, как правило, являются ауксоавтотроф-ными, т. е. не нуждаются в дополнительных факторах роста.
Культуры спорообразующих бактерий, объединяемые в вид Вас. thuringiensis, приобретают в настоящее время особо важное значение в связи с организацией на основе их использования промышленного производства высокоэффективных инсектицидов. Этот вид включает ряд разновидностей, отличающихся друг от друга морфо лого-физиологическими особенностями, а также энтомопатогенной активностью к насекомым. В настоящее время общепринятой является внутривидовая дифференциация Вас. thuringiensis на серотипы по сероагглютинации к жгутиковому антигену отдельных культур бактерий. Подобное разграничение проводится и по многим другим отличительным особенностям бактерий. Некоторые разновидности этого вида образуют энтомоцидный токсин нуклеотидной природы, который, в отличие от кристалловидного токсина, находится в растворимом состоянии в культуральной жидкости бактерий и не разрушается при кипячении.
При внутривидовом разграничении культур этого вида учитываются и многие другие признаки, обычно используемые для определения микроорганизмов. Энтомопатогенные спорообразующие бактерии являются возбудителями болезней тутового шелкопряда, наносят огромный вред шелководству. Они широко распространены в почве, эпифитной микрофлоре листа шелковицы и обнаруживаются в нормальной микрофлоре гусениц тутового шелкопряда. При благоприятных для развития условиях эти микроорганизмы вызывают гибель гусениц, что в ряде случаев приводит к быстрому распространению инфекции. Вызываемое инфекцией заболевание гусениц проявляется, как правило, в последнем возрасте гусениц или в период окукливания. Значительный ущерб, наносимый шелководству, обусловлен бактериальными болезнями бациллярного происхождения. Энтомопатогенные спорообразующие бактерии — возбудители бактериозов тутового шелкопряда — особенно часто обнаруживаются в южных районах шелководства.
Бактерии вида Вас. thuringiensis, продуцирующие энтомоцидные кристалловидные токсины, в большинстве случаев обитают среди кишечной микрофлоры различных насекомых. Однако обнаружены и определенные эколого-географические ареалы распространения отдельных разновидностей. Приспособленность этих бактерий к среде обитания и условиям существования специфична.
Вредоносность аэробных спорообразующих бактерий и их патогенные свойства
Различные виды спорообразующих бактерий широко распространены в окружающей среде и нередко рассматривались как возбудители разнообразных процессов порчи пищевых продуктов.
Спорообразующие бактерии стали важным объектом исследований в связи с развитием консервной промышленности. Было выполнено много работ по изучению устойчивости и выживаемости бактериальных спор при различных методах консервирования продуктов. Особое внимание уделялось изучению анаэробных видов спорообразующих бактерий, в частности ботулиническому бациллу, который образует ботулинический яд, вызывающий тяжелые пищевые отравления.
Различные виды аэробных спорообразующих бактерий, особенно термофильные формы, вызывают порчу консервированных продуктов, например сгущенного молока, а также различных кондитерских изделий. Вас. coagulans, особенно его термофильные формы, обладающие способностью развиваться в кислой среде, вызывают порчу томатов.
Весьма распространенными формами спорообразующих бактерий, вызывающих порчу консервированных продуктов, являются также бактерии группы сенного и картофельного бацилла. Эти бактерии широко распространены в почве и воздухе, постоянно загрязняют пищевые продукты, а также материалы и сырье, используемые для их выработки. Описаны случаи порчи консервов с бомбажем банок, при которых отмечалось массовое распространение бактерий группы сенного бацилла. По наблюдениям многих авторов, процент порчи консервированных продуктов находится в прямой зависимости от обсемененности их споро-образующими бактериями перед стерилизацией,особенно часто это обнаруживается в овощных консервах, загрязненных почвенной микрофлорой.
При микробиологических исследованиях ряда консервированных продуктов, подвергшихся порче, обнаружено значительное количество спороносных бактерий группы сенного и картофельного бацилла. Среди них нередко преобладают солестойкие (галофильные) и кислотоустойчивые формы с выраженными ферментативными свойствами.
При порче пищевых продуктов происходит ферментативный распад белковых и липидных компонентов с образованием вредных соединений. Поэтому использование в пищевой и консервной промышленности некоторых добавок, например агара и желатина, способствует в ряде случаев порче ряда кулинарных изделий и продуктов.
Аэробные спорообразующие бактерии неоднократно являлись причиной порчи консервируемых продуктов крови. Отмечены случаи смертельных исходов при использовании испорченной консервированной крови и плазмы главным образом бактериями из группы сенного и картофельного бацилла. В этих случаях основная роль в порче подобных продуктов принадлежит таким ферментам, как гемолизины.
Распространенными возбудителями порчи различных пищевых и кулинарных изделий, а также молочных продуктов являются часто встречающиеся в природе спорообразующие бактерии, объединяемые в вид Вас. cereus. В литературе описаны случаи пищевых отравлений, вызванных этими бактериями, которые, как правило, не смертельны. Пищевые расстройства и токсические явления, развивающиеся после потребления в пищу подвергшихся порче продуктов, связывают с действием ферментов группы фосфолипаз.
Бактерии этого вида постоянно обнаруживаются при анализах пищевых продуктов, особенно полуфабрикатов и готовых кулинарных изделий. Источником загрязнений микроорганизмами являются воздух и различные предметы окружающей среды. При хранении пищевых продуктов в отсутствие холода всегда создаются благоприятные условия для развития и массового размножения бактерий.
Среди аэробных спорообразующих бактерий типично патогенным для человека и животных является только сибиреязвенный бацилл. Остальные виды этих бактерий вирулентными свойствами для теплокровных, по-видимому, не обладают. Имеющиеся сообщения о выделении различных видов спороносных бактерий при разнообразных инфекционных поражениях или болезнях не могут служить достоверным основанием для заключений о патогенности этих бактерий для животных и человека.
Вызываемые культурами некоторых видов спороносных бактерий воспалительные поражения могут рассматриваться как результат неспецифического действия активного комплекса ферментов. Энтомопатогенные штаммы данной группы бактерий также практически безвредны для человека и животных. Бактерии группы сенного и картофельного бацилла являются возбудителями болезней печеного хлеба и макарон.
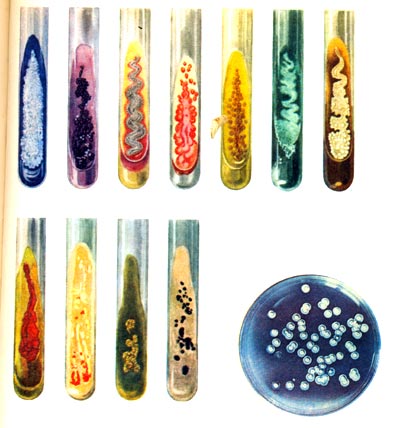
Таблица 40. Пигментация у различных видов актиномицетов в пробирках и чашке Петри.
Разрушение структуры хлеба и разложение содержащихся в нем веществ связано с продуцированием бактериями группы сенной и картофельной палочки активных протеолитиче-ских и амилолитических ферментов. Одним из основных факторов, обусловливающих развитие поражений хлеба, является накопление в зерне и муке спороносных бактерий группы картофельного бацилла. Спороносным бактериям, особенно их термофильным формам, отводится значительная роль в процессах самосогревания зерна.
Некоторые виды спорообразующих бактерий, обладающие выраженными протопекти-назными и протеолитическими свойствами, обладают фитопатогенным действием. Ткани пораженных растений и плодов подвергаются мацерации, болезнь выражается в побурении или загнивании.

Таблица 41. Действие антибиотика актиномицетного происхождения на непарного шелкопряда: 1—лиственница, не пораженная шелкопрядом; 2, 3 — лиственницы, пораженные шелкопрядом; 4 — дубовые ветки, обработанные антибиотиком и зараженные шелкопрядом; 5 — дубовые ветки (контроль); 6 — бабочка шелкопряда.
Большинство описанных фитопатогенных культур относится к группе картофельной бациллы. Эта бактерия поражает различные растения: лен, тыкву, кукурузу, свеклу, плоды апельсина, абрикоса, кабачков и других растений, клубни картофеля, семенники капусты, коробочки хлопчатника. Показано, что картофельный бацилл может вызвать бактериоз початков кукурузы, при искусственном заражении ими бактериями происходило типичное побурение сосудистого пучка кок-сагыза. Из загнивших плодов томатов выделен штамм спорообразующей бактерии, по-видимому, родственной культурам сенного бацилла.
Другой группой спорообразующих бактерий, вызывающих поражения растений, являются Вас. macerans, Вас. polymyxa. Родственный этой группе вид вызывает бактериоз льна. Описаны поражения картофеля, моркови, лука, огурцов и других растений и плодов, вызываемые этими бактериями. Ведущим фактором в этиопатогенезе этих поражений являются ферменты, разлагающие пектин и протопектин.
Применение бактерий рода Bacillus для борьбы с вредоносными насекомыми
Микробиологический метод борьбы с вредоносными насекомыми, именуемый сокращенно микробиометодом, основан на использовании энтомопатогенных микроорганизмов, которые являются возбудителями болезней насекомых или образуют вещества, обладающие энтомоцидным действием. Как правило, микробиометод предполагает использование препаратов живых микроорганизмов. Энтомоцидные продукты жизнедеятельности микроорганизмов не получили пока широкого практического применения.
Современное промышленное производство препаратов микробных инсектицидов основано на использовании в основном двух видов энтомопатогенных бактерий — Вас. thuringiensis и Вас. popilliae.
В 1874 г. Луи Пастер предложил использовать энтомопатогенные бактерии для борьбы с филлоксерой. И. И. Мечников в 1879 г. описал возбудителей бактериальной и грибной болезней хлебного жука и провел успешные опыты по использованию их против этого вредителя. Развивая эти исследования, И. М. Красильщик организовал в 80-х годах прошлого столетия лабораторию по производству и применению микробов, обладавших энтомопатогенным действием; ученый установил губительное действие некоторых микробов на хлебного жука и ряд других вредителей.
В последующие годы были описаны возбудители многих болезней насекомых, некоторые культуры энтомопатогенных микробов использовались с переменным успехом для борьбы с вредоносными насекомыми в широких масштабах. Широкий отклик получили работы Д' Эрелля (1911) по использованию выделенной им культуры неспороносной бактерии для борьбы с саранчой в некоторых странах Южной Америки и Северной Африки.
Для развития микробиометода особенно ценными явились работы С. Метальникова, В. Шорина и К. Туманова. В 1922 —1942 гг. ими было выделено и изучено большое число энтомопатогенных форм спорообразующих бактерий, на основе которых было организовано производство инсектицидных препаратов для борьбы с опасными вредителями кукурузы, винограда и хлопчатника.
В 1941 г. в США, используя культуры возбудителей так называемых молочных болезней насекомых, Датки разработал и успешно применил метод получения нового инсектицидного препарата для борьбы с японским жуком.
В 1949 г. Е. В. Талалаев разработал и внедрил в широкую практику метод борьбы с сибирским шелкопрядом с помощью культуры спорообразующей бактерии, принадлежащей к виду Вас. thuringiensis. Открытие в 1953 г. Хэннейем энтомоцидной природы кристалловидных включений у бактерий этого вида положило начало широким исследованиям по использованию его при получении новых эффективных инсектицидных препаратов. В 1959—1960 гг. в СССР, США, Франции было организовано промышленное производство специальных препаратов бактериальных инсектицидов из культур разновидностей Вас. thuringiensis. Микробные и в первую очередь бактериальные инсектициды прочно входят в систему мероприятий по защите растений и широко используются в борьбе со многими вредителями сельского хозяйства.
На основе использования культуры новой разновидности Вас. thuringiensis была разработана технология получения препарата бактериального инсектицида-энтобактерина, промышленное изготовление которого в нашей стране осуществляется с 1959 г. В настоящее время энтобактерин широко применяется для борьбы со многими видами вредоносных насекомых.
В настоящее время основное внимание уделяется получению высокоэффективных культур энтомопатогенных микроорганизмов. Большое значение имеет выявление природы энто-моцидных веществ из микроорганизмов, изучение их строения, механизмов биосинтеза и специфики действия на насекомых. Особую группу энтомопатогенных бактерий составляют виды, продуцирующие энтомоцид-ные токсины при развитии на обычных питательных средах. Наибольший интерес из этой группы представляют аэробные спорообра-зующие бактерии, объединяемые в вид Вас. thuringiensis. Использование бактерий этого вида составляет основу современной промышленности, выпускающей микробные инсектициды.
Характерной особенностью этих бактерий Вас. thuringiensis является их способность продуцировать специфические кристалловидные токсины, обладающие высокой энтомоцидной активностью. Кристаллы имеют ромбовидную (тетрагональную) форму, обнаружива- ются внутри клеток в процессе споруляции с помощью обычной микроскопии живых препаратов бактерий, и их часто именуют как па-распоральные включения или эндотоксины. После завершения процесса образования спор токсины выделяются в среду и обнаруживаются в свободном виде. Величина кристаллов варьирует в зависимости от вида культур бактерий в пределах 1—3 мкм. Энтомоцидные кристаллы — вещества белковой природы, в их состав входят 18 аминокислот. Энтомоцидный кристалл — термолабильный белок, разрушается при температуре около 60° С. Энтомоцидные кристаллы токсичны для чешуекрылых насекомых лишь при пероральном введении: вызывают паралич средней кишки гусениц. Насекомые гибнут при обработке их растворами токсинов очень малой концентрации — введение 1 : 106 и более.
Неоспоримым преимуществом этих препаратов является отсутствие токсического действия на энтомофаги и полезную энтомофауну. Так, они безвредны для медоносной пчелы. Препараты кристалловидных токсинов безвредны для теплокровных животных, рыб и растений. Кроме кристалловидных токсинов, бактерии некоторых разновидностей Вас. thuringiensis выделяют в среду растворимые в воде Энтомоцидные вещества, из которых наибольший интерес представляет термостабильный токсин, или бета-экзотоксин. В отличие от энтомоцид-ных кристаллов это вещество не разрушается даже при 100—120 °С. Оно является сравнительно низкомолекулярным соединением нук-леотидной природы. В составе термостабильного экзотоксина обнаружены два родственных вещества, названных турингиензииами,— А и В. Энтомоцидное действие термостабильного экзотоксина менее специфично — он активен по отношению ко многим видам насекомых. Практический интерес представляет его энтомоцид-ное действие на мух, комаров, совок и некоторых других вредных насекомых. Препараты экзотоксина безвредны для теплокровных животных, рыб и растений.
Микробиологическая промышленность изготовляет инсектицидные препараты из культур некоторых разновидностей Вас. thuringiensis. В нашей стране известны препараты энтобак-терин-3, дендробациллин, инсектин; за границей — биотрол, турицид, агритрол, бактан, ди-пел, спореин и др. Бактериальные инсектициды чаще всего выпускаются в виде порошка. Порошки более эффективны, транспортабельны, удобны для хранения и сохраняют свою активность в течение нескольких лет.
Несомненным преимуществом обладают гранулированные и инкапсулированные формы бактериальных препаратов, с успехом использованные против ряда вредителей. Так, применяются препараты в гранулах с кукурузной мукой, которые эффективны против кукурузного мотылька. Такая форма препаратов позволяет избежать возможное пагубное влияние инсектицидов на полезную энтомофауну.
При производстве инсектицидных препаратов применяются разнообразные добавки (растека-тели, прилипатели или распылители), которые значительно повышают эффективность используемого бактериального инсектицида. Энтомо-цидная активность таких препаратов превышает исходную при одинаковом содержании спор и энтомоцидных кристаллов.
Энтомоцидное действие препаратов из культур бактерий Вас. thuringiensis распространяется в основном на чешуекрылых; число чувствительных к действию Вас. thuringiensis насекомых превышает 150 видов. Насекомые из отрядов перепончатокрылых, двукрылых и жуков нечувствительны к действию этих бактерий. К наиболее вредоносным насекомым, подверженным действию препаратов Вас. thuringiensis, относятся яблонная, черемуховая, плодовая, бересклетовая, магалебская, ореховая, рябиновая, сиреневая, капустная моли; капустная и репная белянки, боярышница, ивовая волнянка, сосновый, сибирский, кольчатый, непарный шелкопряды, златогузка, листовертки, розанная, почковая, всеядная, плодовая, капустная, мельничная огневки, зимняя кустовая, желтоусая пяденица, американская белая бабочка, яблонная плодожорка, вишневый слизистый пилильщик, кукурузный мотылек, мальвовая моль, табачный бражник, коробочный червь хлопчатника. Безвредность бактериальных инсектицидов для растений позволяет применять препараты в любую фазу вегетации растений. Норма расхода препаратов зависит от вида насекомых-вредителей, метеорологических условий, размеров обрабатываемых растений, а также характера опрыскивания. Для порошковидных препаратов энтобак-терина установлены следующие дозировки: обработка овощных культур — 1,0—3,0 кг на 1 га, садовых и других культур — 3,0—5,0 кг на 1 га.
Перспективно совместное использование бактериальных инсектицидов с различными химическими средствами. При этом отмечается синергизм их энтомоцидного действия; спектр энтомоцидного действия препаратов расширяется, уменьшаются нормы их расхода.
Иммунитет и привыкание к действию энтомоцидных кристаллов и других бактериальных токсинов у чувствительных видов насекомых не отмечаются. При проникновении в кишечник насекомых достаточных доз этих соединений отмечается 100-процентная гибель вредителей.
В природе основным условием проявления вирулентных свойств энтомопатогенных микроорганизмов является массовое развитие бактерий в кишечнике чувствительных видов насекомых. Это развитие происходит во всех случаях нарушения физиологии насекомых, неблагоприятного воздействия на них внешней среды и наличия в кишечнике условий, способствующих развитию этих бактерий.
Исключительно перспективны для производства инсектицидных препаратов культуры возбудителей молочных болезней, в особенности Вас. popilliae. Культуры этого вида — очень эффективное средство борьбы с японским жуком — опаснейшим вредителем, поражающим около 300 видов растений. Бактерии выращивают непосредственно в организме личинок японского жука. Погибших, высушенных личинок вносят в почву, где споры бактерий долгое время остаются жизнеспособными и вирулентными для жука. Выращивание Вас. popilliae в организме насекомых — весьма трудоемкий процесс. В последние годы достигнут значительный успех в изучении физиологических и биохимических особенностей культур этой группы бактерий, что послужит основой для поисков способов выращивания их на искусственных питательных средах.
Методы выращивания Вас. popilliae на искусственных средах позволят получить препараты бактериальных инсектицидов для борьбы с особо вредоносными насекомыми, в том числе с жуками, хрущами и другими видами, против которых практически нет эффективных средств борьбы. Бактериальные инсектицидные препараты качественно отличны от химических инсектицидов. Они не обладают контактным действием, более специфичны и активны, кратность их применения значительно меньше, чем химических средст-в, привыкание к ним насекомых не отмечено. Вместе с тем повышение эффективности при совместном использовании бактериальных и химических инсектицидов служит основанием для разработки единой системы интегрированной борьбы с вредоносными насекомыми.
Среди микробных метаболитов, которые могут быть применены в качестве инсектицидных препаратов, определенное значение имеют токсические антибиотики. Выделен ряд антибиотиков (антимицин А, пиерицидин, деструксины и др.), обладающих широким спектром инсектицидного действия.
Порядок хламидобактерии (CHLAMYDOBACTERIALES)
К хламидобактериям относят гетеротрофные бактерии, растущие обычно в виде нитей и образующие чехлы (влагалища), где аккумулируются гидраты окиси железа. Молодые клетки, выходящие из чехла, подвижны, облададают полярным или субтерминальным жгутиком. Микроорганизмы этого порядка широко распространены в природе и бурно развиваются в тех местах, где происходит постоянное загрязнение воды и почвы пищевыми отходами.Семейство хламидобактерии (CHLAMYDOBACTERIACEAE)
Характерными представителями порядка Сhlаmydobacteriales являются организмы, относимые к семейству Chlamydobacteriaceae. Наиболее типичным среди этих организмов является род сферотилюс (Sphaerotilus). Представители этого рода — типичные обитатели сточных вод. Растут в загрязненной проточной воде, в отстойниках сахарных заводов, на плотинах и градирнях, на твердых поверхностях, где образуют наросты, напоминающие густой мех. Трубы, отстойники, сточные канавы, ямы в короткий срок забиваются этими бактериями. Типовой вид — Sphaerotilus natans — грамотрицательная бактерия с полярным жгутиком. Растет в форме нитей, формируя цепочки клеток, окруженных трубчатым чехлом, состоящим из полисахарида (чехол — разновидность капсулы).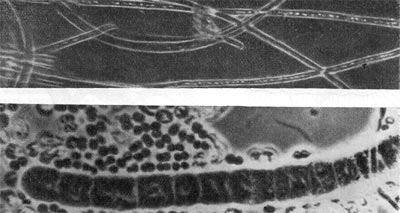
Рис. 66. Многоклеточные бактерии Beggiatoa, выделенные из разных источников (по Броку, 1970).
Клетки размножаются внутри чехла поперечным делением, после чего наружу выходят подвижные формы.
Представители другого рода — лептотрикс (Leptothrix), называемые также «охровыми бактериями», развиваются в воде, богатой железом, в канавах, дренажных трубах и болотах. В местах обитания органические вещества часто связаны с железом (комплексы). Поэтому их чехлы пропитаны железом. Сходной морфологией обладают также нитчатые бесцветные серные бактерии (род Beggiatoa) и представители других родов (Leucothrix, Thiothrix). Они отличаются от других хламидобактерий крупными размерами клеток и способностью окислять минеральные соединения серы, что послужило основанием для выделения этих нитчатых бактерий в порядок Beggiatoales (рис. 66).
Порядок кариофановые (CARYOPHANALES)
В этот порядок М. А. Пешков (1957) выделяет входившие ранее в порядок Chlamydobacteriales гетеротрофные нитчатые бактерии, сходные с сине-зелеными водорослями, но бесцветные. В отличие от хламидобактерий у Caryopha-nales жгутики расположены перитрихиально. Наиболее характерным родом семейства является кариофанон (Caryophanon), впервые описанный М. А. Пешковым. Отдельные клетки этой бактерии обладают жгутиками, и вся многоклеточная особь активно подвижна. Эти грамположительные палочки выделяются обычно из коровьего навоза. Близкие к ней по строению с крупными трихомами (члениками) представители рода осциллоспира (Oscillospira) образуют споры (рис. 67).
Хищные нитчатые бактерии
Крайне своеобразная группа хищных нитчатых бактерий впервые описана на уровне порядка Cyclobacteriales советским микробиологом Б. В. Перфильевым.Клетки этих бактерий постоянно соединены плазмодесмами. Большие группы клеток погружены в слизь и обладают способностью согласованно двигаться. В род диктиобактерий (Dictiobacter) были включены бактерии, образующие микроскопические скопления — бактериальные колонии, состоящие из 100— 200 отдельных довольно мелких клеток (1—6 мкм), связанных плазмодесмами (мостиками). Центральная полость этой группы заполнена гомогенной жидкостью. Во время движения колония захватывает живые микроорганизмы и переваривает их.
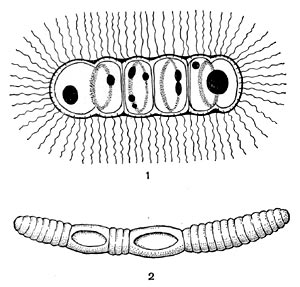
Рис. 67. Схема строения многоклеточных бактерий: 1 — Caryophanon и 2 — Oscillospira (по Пешкову, 1955).
Представители другого рода хищных бактерий — циклобактер (Gyclobacter) представляют собой также многоклеточные колонии палочек. Одной из трех стадий в цикле развития является «сетчатоарканная», когда бактерия охотится, обволакивая жертву «коконом» из клеток и разрушает ее. Третьим родом хищных бактерий является тератобактер (Teratobacter, рис. 68). Б. В. Перфильев наблюдал у этой бактерии ловчее приспособление в виде петель, которое облегчает улавливание жертв, как правило, нитчатых бактерий (в частности, Beggiatoa).
Рис. 68. Устройство ловчего приспособления хищной бактерии Teratobacter (по Перфильеву, Габе, 1961).
Бактерии, подобные описанным, довольно легко обнаружить при постановке простейших опытов. Достаточно внести в колбу с водой небольшое количество богатой органическим веществом почвы или ила, чтобы спустя 10—15 дней на поверхности воды обнаружить микроскопления клеток, соединенных плазмо-десмами в большие группы. Такая форма роста известна также под названием бактодерм, и ради объективности следует отметить, что убедительных доказательств в пользу хищной природы подобных скоплений (микроколоний) все еще недостаточно. Само существование таких многоклеточных агрегатов не вызывает сомнения и является формой существования в природе обычных сапрофитных бактерий.
Другим примером сложных нитчатых объединений клеток могут служить обнаруженные В. И. Дудо и (1972) анаэробные неспорообразующие бактерии, формирующие сложно организованные колонии, состоящие из клеток, упорядоченных в нити, взаимно переплетенных. На почвенных частицах, взятых в качестве питательной среды, эти бактерии образуют воздушные колонии, напоминающие колонии акти-номицетов. При просмотре в сканирующем электронном микроскопе (микроскоп, работающий по принципу отраженного луча) видна сетчатая структура колоний (табл. 38). Отдельные клетки соединены между собой с помощью перетяжек. Из-за задержки в делении клеток перетяжки сохраняются долго. Некоторые колонии таких организмов имеют вид белого пушка, другие — окрашены. Они образованы клетками разных размеров. Эти организмы могут расти на поверхности стекла и минералов в камерах, насыщенных парами воды. Возможно, такие микроколонии способны активно адсорбировать пары воды и запасать впрок, так как огромная «ворсистая» поверхность этих колоний вполне соответствует такой задаче. Большинство этих организмов могут развиваться на почвенных средах (агаризованная почва) с добавлением витаминов и других факторов роста.
Хищные бактерии отражают экологически адекватную (соответствующую среде обитания), но не обязательную форму роста. По-видимому, близки к этой группе и анаэробные нитчатые бактерии.
Порядок железобактерии(FERRIBACTERIALES)
Железо необходимо всем живым организмам. В природе оно существует в органических и неорганических соединениях. Главную роль в круговороте железа в природе играют микробы.
Эти процессы идут по двум каналам: 1) минерализация органических соединений, содержащих железо, при участии гетеротрофных микроорганизмов; 2) окисление восстановленных (закисных) и восстановленных окисных соединений железа.
Минерализацию железосодержащих органических веществ проводят многочисленные гетеротрофные организмы (бактерии, грибы, акти-номицеты). Только специфические возбудители — хемолитоавтотрофы — способны осуществлять второй процесс. Это — представители рода Thiobacillus — грамотрицательных аэробных бактерий. Основной процесс, проводимый ими, описывается следующей схемой: 4Fe2+++4H++02 -> 4Fe3++2H20 Для одних кислотоустойчивых бактерий (выдерживают значения рН, равные 2,5) способность к хемолитотрофному образу жизни (получению энергии за счет окисления ионов за-кисного железа) убедительно доказана. Таким организмом является представитель тионовых бактерий — Thiobacillus ferrooxidans. Для других «классических» железобактерий (например, Gallionella ferruginea) таких данных нет. Возникает сомнение, являются ли они истинными железобактериями.
Рис. 69. Бактерии со слизистыми стебельками: 1 — Nevskia, 2 — Gallionella.
Порядок железобактерий объединяет сборную группу одноклеточных бактерий, способных аккумулировать соединения железа и марганца в результате гетеротрофных процессов. Другие организмы, способные к окислению и восстановлению соединений железа, отнесены к другим порядкам: серобактериям (род Thiobacillus) и нитчатым бактериям (род Leptothrix). Бактерии, отнесенные к порядку железобактерий, разделены на 2 семейства. Многие представители этих семейств обладают своеобразной морфологией и сложным жизненным циклом.
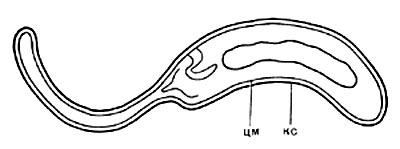
Рис. 70. Схема строения типичной клетки стебельковой бактерии. КС — клеточная стенка, ЦМ — цитоплазматическая мембрана
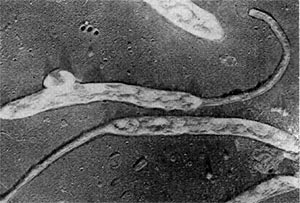 Рис. 71. Типичные клетки бактерий рода Caulobacter. Электронная микрофотография. Увел. X 20 000.
Рис. 71. Типичные клетки бактерий рода Caulobacter. Электронная микрофотография. Увел. X 20 000.Семейство железобактерии (FERRIBACTERIACEAE)
Клетки представителей семейства обладают либо ложными слизистыми придатками, либо истинными стебельками — выростами цитоплазмы. Они широко распространены в природе, и прежде всего в иле и воде пресных водоемов. Семейство представлено 6 родами.
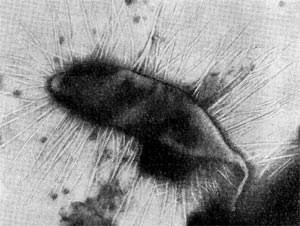 Рис. 72. Стебельковая бактерия с нетипичным тонким стебельком. Увел. X 25 000.
Рис. 72. Стебельковая бактерия с нетипичным тонким стебельком. Увел. X 25 000.Роды галлионелла и невския (Gallionella и Nevskia)
Бактерии, принадлежащие к родам Gallionella и Nevskia (рис. 69), обладают придатками, состоящими из слизи и не связанными с цитоплазмой клетки. Форма и размеры таких придатков сильно варьируют. При исследованиях природных субстратов у бактерий из рода Gallionella легко обнаруживаются почковидные или палочковидные клетки, которые формируют длинные перевитые слизистые стебельки, пропитанные окислами железа. Согласно представлениям, развиваемым Г. А. Заварзиным, природа стебельков Gallionella иная. Стебельки представляют собой скопление живых организмов, не обладающих клеточной стенкой и способных формировать длинные нити. Эти организмы паразитируют на клетках истинных бактерий. Поэтому то, что принималось обычно за бобовидную клетку Gallionella, является лишь клеткой-хозяином, который может быть в разных случаях разным. Такими паразитами могут быть прежде всего микоплазмы, которые будут рассмотрены в соответствующем разделе. Истинные стебельковые бактерии имеют стебелек, сформированный клеточной стенкой и цитоплазматической мембраной. Они отнесены к семейству Caulobacteriaceae.
Семейство сидерокапсы (SIDEROCAPSACEAE)
Все организмы, объединенные в семействе Siderocapsaceae, сходны между собой, являясь, по-видимому, разными экологическими формами одной или нескольких близких бактерий. Известны успешные попытки и описания указанных микроорганизмов в составе одного рода. В это семейство входят палочковидные или кокковидные (часто овальные клетки) неспоро-образующие гетеротрофные бактерии, образующие слизистую капсулу, пропитанную солями железа или марганца. Бактерии, принадлежащие к роду Siderocapsa, имеют небольшие клетки (1 — 2 мкм в диаметре), объединенные в первичные капсулы (по 2—60 и более клеток). Эти капсулы с клетками (общий диаметр 10—20 мкм) объединяются в более сложные агрегаты, где и откладывается железо или марганец. Род Sideromonas объединяет палочковидные бактерии (длина клетки 2 мкм), имеющие капсулы и формирующие группы (пары, цепочки) и скопления. К семейству Siderocapsa-сеае были отнесены описанные в разное время микроорганизмы: род Siderosphaera (объединены по 2 клетки в капсуле), род Sideronema (крупные палочки диаметром 5,0—6,5 мкм, соединенные в цепочки и заключенные в капсулы). Три известных в литературе рода — Naumaniella, Ochrobium, Siderococcus — объединяют, подобные описанным выше, мелкие (диаметр клетки 2 мкм) палочки, не обладающие капсулами. Отложение окислов железа и марганца происходит прямо на клетках.Стебельковые бактерии
Стебельковые бактерии в настоящее время рассматриваются как группа организмов, близкая к эубактериям, и выделяются в семейство кауло-бактерии (Caulobacteriaceae). Принадлежность к определенному порядку не уточнена.
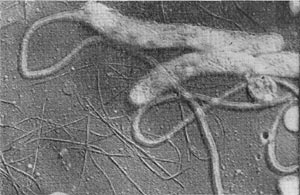 Рис. 73. Клетки стебельковой бактерии пз рода Asticcacaulis. Электронная микрофотография. Увел. X 20 000.
Рис. 73. Клетки стебельковой бактерии пз рода Asticcacaulis. Электронная микрофотография. Увел. X 20 000.Род каулобактер (Caulobacter)
Большинство истинных стебельковых бактерий относятся к этому роду. Похожие на псевдомонады клетки, с одним полярно расположенным жгутиком, прикрепляются тем концом, на котором находится жгутик, к какой-либо поверхности (часто к другой клетке) и формируют стебелек. Стебелек имеет довольно сложное строение. Наружная поверхность образована клеточной стенкой, за которой следует цитоплазматическая мембрана, т. е. стебелек является выростом (выпячиванием) клетки, сохраняющим ее внутреннюю структуру (рис. 70, 71). Внутри стебелька находится много мембранных структур, играющих важную роль в жизнедеятельности клетки. Форма и размеры стебельков сильно варьируют. Встречаются бактерии с очень тонкими стебельками (рис. 72), и даже у одного и того же вида бактерии в зависимости от условий среды могут быть очень длинные (5 —10 мкм и более) изогнутые стебельки и короткие, едва заметные. В почве нередко можно обнаружить клетки стебельковых бактерий, усеянные по всему периметру, включая стебелек, выростами клеточной стенки — фимбриями (рис. 72).
Функции фимбрий у стебельковых бактерий неясны
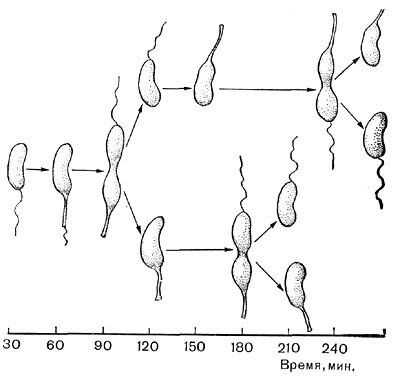 Рис. 74. Стадии жизненного цикла Caulobacter. Схема (по Броку, 1970).
Рис. 74. Стадии жизненного цикла Caulobacter. Схема (по Броку, 1970).
Род стебельковых бактерий — астикакаулис (Asticcacaulis)
Другой род стебельковых бактерий — астикакаулис (Asticcacaulis) — объединяет бактерии, отличающиеся от описанных лишь асимметричным расположением стебелька (рис. 73). Представители этого рода широко распространены в воде пресных и соленых водоемов. Встречаются в почвах. Клетки стебельковых бактерий размножаются поперечным делением, причем дочерняя клетка несет жгутик, но не обладает стебельком, который формируется позже, продолжая нести на дистальном конце жгутик. Это придает большое своеобразие циклу развития этих бактерий, что хорошо видно из схемы (рис. 74).Следует отметить, что на конце стебелька часто образуется небольшая слизистая подушечка, обеспечивающая плотное прикрепление к поверхностям. Отмечены случаи прикрепления стебельковых бактерий к живым клеткам других бактерий, что породило предположения об их паразитизме, оказавшиеся неверными. Стебельковые бактерии выступают здесь в роли эпибионтов — организмов, поселяющихся на других организмах или рядом, но не связанных симбиотическими или иными взаимоотношениями.
Стебельковые бактерии — типичные обитатели пресных водоемов, илов и почв. Будучи гетеротрофами, они развиваются в природе при низких концентрациях органических веществ. В лабораторных условиях их почти всегда можно обнаружить в виде пленки даже на поверхности дистиллированной воды. В этих условиях они развиваются, используя растворяющиеся в воде питательные вещества, поступающие из воздуха. Вполне вероятно, что они способны также усваивать летучие органические соединения, содержащиеся в воздухе. Сведения о возможности воздушного питания бактерий были получены несколько десятков лет назад видным советским ученым Н. Г. Холодным. Эти бактерии в большей мере, чем другие гетеротрофы, обладают способностью к гетеротрофной фиксации углекислоты из воздуха, т. е. включением С02 в клетку непосредственно без энергетических затрат за счет фотосинтеза или энергии, получаемой при превращении неорганических веществ.
Роль стебельковых бактерий в природе определяется их физиологическими особенностями как гетеротрофных микроорганизмов, способных развиваться в зонах обеднения, где неактивны более требовательные к пище сапрофиты.
Порядок почкующиеся бактерии (HYPHOMICROBIALES)
К порядку почкующихся бактерий (Hypho-microbiales) относятся гетеротрофные грамотрицательные неспорообразующие бактерии со своеобразной морфологией. Их характерной особенностью является наличие гиф — выростов, образованных клеточной стенкой и цито-плазматической мембраной, и размножение путем образования почек на гифах. Многие виды способны аккумулировать окислы железа и марганца. Водные почкующиеся бактерии часто встречаются в больших количествах в олиготрофных озерах и нередко в почвах при сильном увлажнении. Рост почкующихся бактерий задерживается даже при небольших концентрациях аминокислот и углеводов. Тем не менее они малочувствительны к повышенным концентрациям нитритов и минеральных солей. В порядок входит семейство Hyphomicrobiaceae.
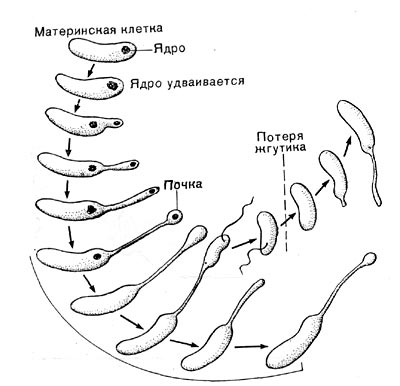 Рис. 75. Стадии жизненного цикла Hyphomicrobium. Схема (по Броку, 1970).
Рис. 75. Стадии жизненного цикла Hyphomicrobium. Схема (по Броку, 1970).Род гифомикробиум (Hyphomicrobium)
Представители этого рода имеют вид вытянутых палочек (0,5 X 2 мкм), от одного из полюсов которых отходит гифа. Весьма своеобразным оказывается цикл развития этих организмов. Вначале материнская клетка, прикрепленная к субстрату, образует тонкий вырост, который удлиняется, формируя гифоподобную структуру. На конце гифы образуется почка. Почка разрастается, и формируется жгутик. Затем дочерняя клетка отделяется от материнской и уплывает. Потеряв жгутик и завершив созревание, дочерняя клетка формирует гифу и почку. Почки могут образоваться вновь на материнской клетке. Возможны разные варианты такого цикла (рис. 75). В некоторых случаях дочерние клетки не отделяются от материнских и образуют гифы и почки на другом полюсе клетки. Возникает сложное скопление гиф и клеток. Иногда почки образуются на материнской клетке без гиф. В других случаях гифы возникают на обоих полюсах клетки (рис. 76). В отличие от стебельковых бактерий прикрепительный матеррил образуется у почкующихся бактерий на полюсе, обратном по отношению к гифе. При этом нередко образуются упорядоченные скопления клеток — розетки.
Следует отметить, что размножение почкующихся бактерий начинается делением ядра. Одно из ядер по гифе переходит в почку, которая, достигнув величины клетки, отделяется. Почкующиеся бактерии сильно изменчивы и в зависимости от условий среды меняют морфологию. Развитие часто бывает усложненным. Одни виды развиваются по типу других. Часто образуются сидячие (долго растущие) почки. Последовательность роста гифы и почек случайна (рис. 77).
Род педомикробиум (Pedomicrobium)
К этому роду почкующихся бактерий, описанному Т. В. Аристовской (1961), относят микро-аэрофильные бактерии, аккумулирующие железо и марганец. Они имеют вид крупных овальных, реже палочковидных клеток, образующих от одной до четырех ветвящихся нитей. Эти нити, как правило, заканчиваются почками, которые представляют собой дочерний организм. Некоторые виды (Pedomicrobium podso-licum) обладают мелкими клетками (диаметр 0,5—1,5 мкм), соединяющие их нити чрезвычайно тонки. Иногда нити сильно редуцированы, и в результате почкования образуются грозди кокков разного размера. В лабораторных условиях не удается выделить культуры этих бактерий в чистом виде, а в случае успешного выделения некоторых штаммов культура погибает.
 Рис. 76. Клетка Hyphomicrobium с двумя стебельками. Электронная микрофотография. Увел. X 20 000.
Рис. 76. Клетка Hyphomicrobium с двумя стебельками. Электронная микрофотография. Увел. X 20 000.Род анкаломикробиум (Ancalomicrobium). Сюда относятся одноклеточные бактерии, имеющие от 2 до 8 отростков длиной до 3 мкм, отходящих от клеток. Отростки могут раздваиваться, но сами не почкуются (рис. 78—80). Почки образуются на клетке. Клетки неподвижны, ведут прикрепленный образ жизни.
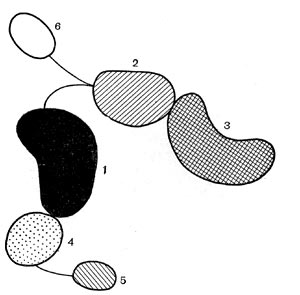 Рис. 77. Последовательность роста гиф и почек Hyphomicrobium. (Цифры означают поэтапность перехода к росту разных участков гнф и почек.) Схема.
Рис. 77. Последовательность роста гиф и почек Hyphomicrobium. (Цифры означают поэтапность перехода к росту разных участков гнф и почек.) Схема.Род простекомикробиум (Prosthecomicrobium)
Микроорганизмы, относящиеся к этому роду, сходны с представителями рода Prosthecomicrobium, но с более короткими (1 мкм) отростками, отходящими от клетки в стороны (рис. 81, 82). Клетки делятся двойной перетяжкой. Молодые клетки могут передвигаться, имея полярный жгутик. Взрослые клетки обладают способностью прикрепляться к субстрату. Имеются пигментированные виды. Представители обоих описанных выше родов широко распространены в воде, илах и во многих почвах. Подобно стебельковым бактериям, они являются типичными представителями экологической группы олиготрофных бактерий.
 Рис. 78. Клетка с гифами (простеками), близкая по морфологии к организмам из рода Ancalomicrobium. Электронная микрофотография. Увел. X 20 000.
Рис. 78. Клетка с гифами (простеками), близкая по морфологии к организмам из рода Ancalomicrobium. Электронная микрофотография. Увел. X 20 000.Другие представители почкующихся бактерий
К почкующимся бактериям относят также ряд других организмов.
Род бластобактер (Blastobacter)
Род бластобактер (Blastobacter) представлен видами, наблюдавшимися в воде пресных водоемов. Обычно слегка изогнутые клиновидные клетки собраны в плотные розетки. На свободных концах клеток образуются почки.
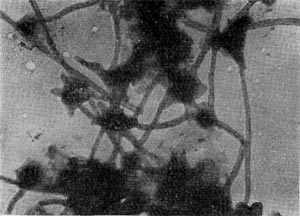 Рис. 79. Скопление клеток из рода Ancalomicrobium Суспензия почвы в электронном микроскопе. Увел. X 15 000.
Рис. 79. Скопление клеток из рода Ancalomicrobium Суспензия почвы в электронном микроскопе. Увел. X 15 000.Давно известный микроорганизм, обнаруженный еще С. Н. Виноградским более полувека назад, — нитробактер (Nitrobacter) — был впоследствии отнесен к почкующимся бактериям. Нитробактер способен окислять нитрит до нитрата, играя большую роль в круговороте азота в природе. К почкующимся бактериям относят и ряд своеобразных микробов, обнаруженных впервые Б. В. Перфильевым при исследовании пресных озер. Это гетеротрофные микроорганизмы, аккумулирующие в колониях железо и марганец: металлогениум (Metallogenium), кузнецовиа (Kuznezovia). Эти организмы, по-видимому, ответственны за образование озерных руд. Типичная стадия развития Metallogenium — микроколония, в форме паучка, составлена радиально расходящимися нитями, покрытыми окислениями марганца. После растворения окислов марганца удается часто обнаружить мелкие почкующиеся клетки, связанные плазматическими нитями. На нити вырастает короткая ножка, на которой образуется почка. Почка прорастает и вновь возникает паукообразная микроколония.
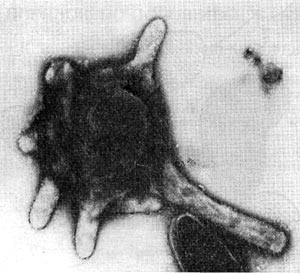 Рис. 80. Молодая клетка Ancalomicrobium. Электронная микрофотография. Увел. X 30 000.
Рис. 80. Молодая клетка Ancalomicrobium. Электронная микрофотография. Увел. X 30 000.Микроколонии организмов из рода Kuznezovia несколько иные, чем у Metallogenium, но по природе эти организмы сходны. За последние годы проведены расследования, позволившие предположить иную природу Metallogenium, Kuznezovia и других подобных организмов. Получены доказательства в пользу того, что эти организмы являются сапрофитными микоплазмами (см. стр. 311). Возможно также, что они паразитируют на других организмах (грибах). При росте на специфических для микоплазм средах можно видеть образование большого числа очень мелких телец, которые проходят через бактериальные фильтры и на средах для Metallogenium дают три-хосферический рост (паучки) с осаждением окислов марганца возле микроколонии. Аккумуляция марганца, естественно, не имеет ничего общего с автотрофией и является следствием тех же процессов, которые были описаны для некоторых неспороносных бактерий. Имеются и другие мнения, что свидетельствует о незавершенности исследований по выяснению природы Metallogenium.
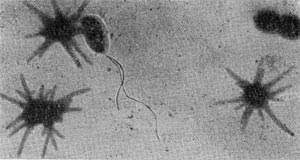 Рис. 82. Клетка неизвестного микроорганизма, напоминающего Prosthecomicrobium. Увел. X 25 000.
Рис. 82. Клетка неизвестного микроорганизма, напоминающего Prosthecomicrobium. Увел. X 25 000.Лучистые грибки. Порядки актиномицеты (ACT1NOMYCETALES)и актинопланы(ACTINOPLANALES)
Лучистые грибки известны под общим названием Actinomyces — актиномицеты. Они давно привлекают внимание специалистов различных областей микробиологической науки. Еще не так давно работы, посвященные этой группе микроорганизмов, выполнялись преимущественно медиками. Чрезвычайно широкое распространение акти-номицетов в природе дает основание полагать, что этим организмам принадлежит большая роль в круговороте веществ, как органических, так и минеральных. Среди актиномицетов имеются аэробы и анаэробы, мезофилы и термофилы. Актиномицеты преимущественно сапрофиты, но среди них есть и паразиты, приносящие вред людям, животным и растениям. К классу Actinomycetes, кроме типичных актиномицетов, относятся проактиномицеты, микобактерии и микококки. Для всех этих организмов характерно ветвление клеток. Проактиномицеты отличаются от актиномицетов тем, что имеют мицелий только в ранней стадии развития. Молодые нити мицелия вскоре распадаются на палочки и кокки. Проактиномицеты образуют колонии тестообразной консистенции, голые или покрытые налетом воздушного мицелия. Микобактерии можно рассматривать как промежуточную форму между актиномицетами и эубактериями. У микобактерии нет настоящего мицелия, в отдельных случаях наблюдаются нитевидные элементы. Палочковидные клетки микобактерии имеют неправильные очертания, контуры их неровные. В отличие от истинных бактерий клетки микобактерии способны ветвиться. В работах, посвященных бактериям, микококки рассматриваются как генетически однородная группа. Внешне микококки похожи на микрококки. Иногда микококки образуют палочковидные клетки с ветками.
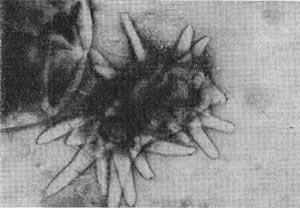 Рис. 82. Клетка неизвестного микроорганизма, напоминающего Prosthecomicrobium. Увел. X 25 000.
Рис. 82. Клетка неизвестного микроорганизма, напоминающего Prosthecomicrobium. Увел. X 25 000.В последние годы лучистые грибки приобрели большое практическое значение в связи с их способностью продуцировать различные физиологически активные вещества, используемые человеком. Из многих культур актиномицетов получают лекарственные препараты — антибиотики, используемые в медицине, ветеринарии, растениеводстве, пищевой промышленности. Актиномицеты способны продуцировать витамины, гормоны, ферменты, токсины, ростовые вещества, аминокислоты и другие полезные для человека биологические вещества. Важную роль актиномицеты играют в процессах почвообразования и формировании плодородия почв. Им приписывают различные функции в оздоровлении почв.
Для того чтобы правильно распознать и определить лучистые грибки, чтобы выявить нужные биологические вещества, вырабатываемые ими, необходимо знать точную характеристику этой группы микроорганизмов. Первые сведения об актиномицетах появились в 1874 г., когда немецкий ученый Ф. Кон выделил и описал микроб с тонкими ветвящимися нитями. В 1878 г. Ц. Гарц обнаружил нитевидные сплетения с лучистым расположением нитей. Найденный микроорганизм был назван им лучистым грибком — Actinomyces.
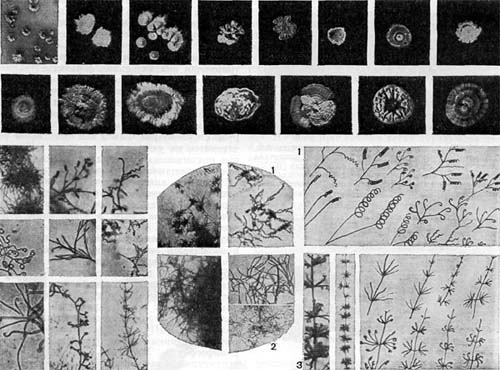 Рис. 83. Вверху — форма колоний актиномицетов на поверхности плотных питательных сред. Внизу - типы спороносцев: 1 — спиральные; 2 — прямые; 3 — мутовчатые; 4 — примитивные.
Рис. 83. Вверху — форма колоний актиномицетов на поверхности плотных питательных сред. Внизу - типы спороносцев: 1 — спиральные; 2 — прямые; 3 — мутовчатые; 4 — примитивные.Первые сведения о строении актиномицетов относятся к началу XX века. Исследования этого периода были сосредоточены главным образом на болезнетворных (патогенных) формах актиномицетов как возбудителях заболеваний человека и животных, вызывающих актиномикозы. В 1921 г. немецкий исследователь Р. Лиске обобщил все накопленные за 50 лет данные о происхождении и свойствах актиномицетов, а также на основании собственных наблюдений описал морфологию различных видов актиномицетов. С 1930 г. исследования актиномицетов проводил Н. А. Кр асильников, который всесторонне изучил эту группу организмов.
Огромное значение имеют работы С. Ваксмана, который вместе с А. Шатцем в 1945 г. выделил из культуры актиномицета антибиотик стрептомицин, нашедший широкое применение в медицине. Это открытие послужило толчком к интенсивному исследованию этой группы микробов и поискам продуцируемых ими новых веществ.
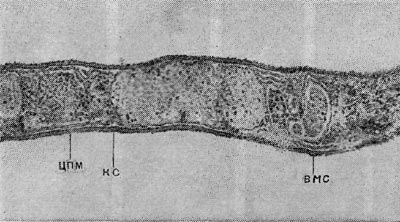 Рис. 84. Ультраструктура клеток актиномицета Act. streptomycini. Окружает клетку плотная клеточная стенка (КС), за ней идет трехслойная мембрана (ЦПМ). Центральная часть клетки заполнена цитоплазмой, в которой хорошо видны рибосомы и мембранные структуры (ВМС). Увел. X 30 000.
Рис. 84. Ультраструктура клеток актиномицета Act. streptomycini. Окружает клетку плотная клеточная стенка (КС), за ней идет трехслойная мембрана (ЦПМ). Центральная часть клетки заполнена цитоплазмой, в которой хорошо видны рибосомы и мембранные структуры (ВМС). Увел. X 30 000.За последнее время выявлены и изучены многие тысячи представителей класса Actinomycetes. Описаны новые формы, которые способны вырабатывать полезные для человека вещества. Была создана антибиотическая промышленность. Лучистые грибки различаются между собой по своему строению и развитию клеток, морфологически, физиологически, культурально, биохимически. Среди них есть организмы со сложным строением плодоносных органов и организмы весьма упрощенного строения. По этому признаку актиномицеты разделяются на две группы — высшие и низшие формы. К высшим относятся организмы с хорошо развитым мицелием и обладающие особыми органами плодоношения, при помощи которых актиномицеты размножаются. В эту группу входит большинство основных семейств и родов, составляющих класс Actinomycetes. К низшим формам относятся организмы, не образующие мицелия и имеющие палочковидные и кокко-видные клетки.
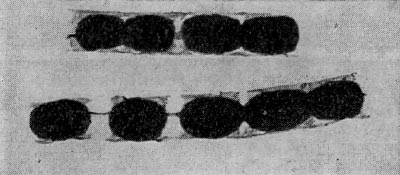 Рис. 85. Фрагментационный способ образования спор.
Рис. 85. Фрагментационный способ образования спор.


