




История происхождения и развития жизни
3.3.3. Что доказывают наблюдаемые последствия отбора?
Наряду с изолированным изучением отдельных факторов эволюции, необходимо также убедиться в том, можно ли соотнести друг с другом существующие научные выводы, если сделать полный обзор всех отраженных в литературе факторов. Так, например, факторы мутации и отбора нельзя рассматривать отдельно друг от друга: под воздействием отбора в генофонде остается лишь малая часть возникших в результате мутаций аллелей (о воздействии селекции см. также рис. 3.15). Если допустить мутации в генофонд без отбора, то это может привести к дегенерации вида, так как мутации для их носителей большей частью вредны. Так отбор "заботится" о сохранении видов.
Существует три различных механизма действия естественного отбора: стабилизирующий, движущий (направленный) и дизруптивный. О стабилизирующем отборе говорят в том случае, если он способствует сохранению определенного состояния и исключает появление мутантов. Достигла, к примеру, оптимальная длина клюва одной популяции птиц в определенном радиусе обитания 15 мм, и отбор стремится "закрепить" эту длину клюва (рис. 3,16а). Обычно на стабилизирующий отбор можно рассчитывать тогда, когда условия окружающей среды остаются неизменными.
Движущий отбор, напротив, происходит в том случае, когда односторонний "селекционный натиск", направленный на изменение одного признака, достиг цели. "В нашем примере трансформирующий отбор имеет место, когда, ввиду изменившихся условий окружающей среды, для популяции птиц стало предпочтительней иметь либо удлиненные, либо укороченные клювы (рис. 3.16b). Изменение признаков можно наблюдать лишь когда наступает изменение условий окружающей среды.
Редкие дизруптивные отклонения закрепляются тогда, когда крайние значения вариационной изменчивости становятся предпочтительными с точки зрения селекции. В нашем примере это означает, что особи с наиболее короткими или длинными клювами имеют наибольшие шансы выжить (рис. 3.16с).
Следствием мутационного процесса является то, что следующие одна за другой мутации, которые должны положить начало возникновению эволюционного новообразования, лишь тогда могут стать отправной точкой, когда каждый отдельный мутационный шаг несет в себе какое-либо селективное преимущество или, по крайней мере, не связан ни с каким явным недостатком. Это означает: селективно-вредные промежуточные стадии не могут привести к эволюционному возникновению новой структуры или нового органа. Г. Оше демонстрирует это наглядно, когда утверждает, что просто перестройкой не может ограничиваться развитие организмов. Каждая отдельная стадия эволюции должна улучшать способность к выживанию (см. разделы 3.3.6 и 4.3).
Здесь следует привести еще одно короткое замечание о механизме отбора: если говорят о том, что селекция "обуславливает", "способствует", то это не следует понимать так, что селекция есть действующий субъект! Речь идет лишь об образе выражения безличных и нецеленаправленных в своем воздействии условий окружающей среды.
В дальнейшем на ряде примеров мы рассмотрим вопрос, какую часть эволюционных процессов можно наблюдать в природе или лаборатории: что доказывают наблюдаемые селекционные процессы? Затем мы рассмотрим, в какой взаимосвязи находятся процессы приспособляемости и развития от простого к сложному.
Бабочки березовой пяденицы (Biston betularia)
Преимущественное распространение темных (меланиновых) форм березовой пяденицы относительно светлых форм считается наглядным примером наблюдаемой эволюции. Распространено даже мнение, что именно в этом случае разыгрывается "великолепное действо", в котором можно "наблюдать эволюцию". Наблюдения за березовыми пяденицами в Англии ДАЙЛ (1980, с. 93) описывает следующим образом (см. также рис. 3.17):
"В 1848 году впервые был найден экземпляр черной окраски в районе Манчестера (средняя Англия). В последующие десятилетия наблюдалось все возрастающее число меланиновых особей. В 1895 г. было проведено количественное исследование, которое показало, что в районе Манчестера 95% всех березовых пядениц были черного цвета. Сегодня 100% светлоокрашенных особей можно встретить лишь на отдаленном севере, западе и юге Британских островов. Подобная тенденция к потемнению окраски наблюдается, между тем. у 70 видов бабочек в Англии и других, индустриально развитых странах. Этот феномен имеет место исключительно у тех видов, для которых привычным местом обитания является открытая поверхность".
Но из этого описания нельзя, тем не менее, сделать вывод, что произошла эволюция. Во-первых, не до конца выяснено, откуда взялись первые темные формы: то ли в результате новых мутаций, то ли темные формы раньше не были открыты. К тому же, если исходить из того, что действительно имело место мутационное происхождение, то речь может идти только о микроэволюции, так как и светлые формы имеют темно-коричневое красящее вещество - меланин, принимающий участие в окраске меланиновых форм (светлые формы имеют коричневые пятна или коричневую кайму на крыльях), так что потемнение могло происходить и без образования новых структур. На самом деле причина кроется в изменении генетически обусловленного процесса синтеза меланина.
Распространение и изменение светлых форм не имеет ничего общего с макро-эволюцией, так как единственное изменение (наступившее в результате селекционного воздействия) - множество аллелей, отвечающих за окраску бабочек. Происходит явное смещение в частотах аллелей. Пример с березовыми пяденица ми наглядно показывает, что отбор может в основном зависеть от условий окружающей среды и что приспособленность есть качество относительное (зависимое от условий окружающей среды). Значит, это не может служить доказательством описанного в синтетической теории механизма развития от простого к сложному.
Маскировочные окраски и миметизм
Феномен маскировочной окраски и миметизм (маскировка путем подражания, рис. 3.18) тесно смыкается с примером березовых пядениц. Здесь, как и в некоторых других примерах, которые мы будем обсуждать, необходимо совершенно ясно различать явление сохранения и возникновения новых соответствующих структур. На основании стабилизирующего отбора плохо замаскированные особи вымирают; том самым предотвращается потеря маскировочной окраски или эффекта подражания. Как возникли эти качества - это совершенно другой вопрос, который должен остаться открытым, пока не удастся экспериментальным путем вызвать маскировочную окраску у "незамаскированных" форм (при этом сами они должны происходить не от некогда имевших маскировочную окраску форм, так как "маскировочные гены" могут присутствовать в организме в рецессивном состоянии).
Но если бы это и произошло, то нужно было бы еще проверить, являются ли возникшие структуры новыми, например, новыми пигментами. В каждом случае следует провести детальные генетические исследования, прежде чем говорить о возникновении эволюционного новообразования. Более подробно это освещается в разделе 3.3.4 на примере из жизни бактерий.
Мимикрия
Мимикрия (ложное защитное обличье) - это "подражание внешнему виду хорошо защищенного животного другим незащищенным животным из другой систематической группы" (FIOR, 1980, с. 6). К примеру, безобидный шершневый бражник повторяет внешний вид шершня (рис. 3.19). Этот феномен может быть аргументирован так же, как и явление защитной окраски и подражания.
Мы установили:
Сохранение признаки или структуры, с одной стороны, и возникновение новых, с другой - обуславливаются абсолютно разными факторами и механизмами: сохранение объясняется отбором (микроэволюцией, а возникновение происходит путем макроэволюции.
Коэволюция
Феномен так называемой коэволюции имеет сходную природу. Под коэволюцией подразумевается взаимное приспособление двух организмов. Например, "взаимное приспособление" цветка и насекомого рассматривается как результат коэволюции. Известны орхидеи-офрисы (рис. 3.20), которые напоминают по виду мух, пчел и шмелей. Лепестки нижней губы этих орхидей до деталей обманно схожи с насекомыми по форме, цвету, запаху и волосяному покрову, из-за чего последние принимают их за половых партнеров и ищут с ними контакта. При этой попытке пыльца переносится на насекомых, а затем и на следующий цветок.
По эволюционной модели следует предположить, что цветок и насекомое развивались в постоянной взаимозависимости, так что цветы "реагировали" на изменения "посетителей", а "посетители" - на изменение цветов при помощи механизма отбора мутантов, и постепенно возникло это удивительное сходство. В основе этого объяснения лежит умозрительная конструкция, не основанная на результатах наблюдения. Механизм возникновения взаимного уподобления цветка и насекомого путем мутации и отбора не предмет наблюдений, а только вывод синтетической теории, на котором в настоящее время выстраивается доказательство. Синтетическая теория в данном случае ничего не доказывает, но дает предпосылки для размышлений. Единственным наблюдаемым фактом является имеющийся в настоящее время феномен приспособления, а не механизм этой адаптации.
Понятие "коэволюция" является искусственным, которое только уводит от объяснения, а не помогает найти его. До сих пор нельзя объяснить механизм возникновения подобной взаимозависимости генетикой или естественным отбором. Это следовало бы, вероятно, считать задачей одной из наук, базирующихся на опыте (см. раздел 1.1).
Неспособные к полету насекомые с островов Кергелен
Убедительным примером воздействия селекции является появление насекомых, не имеющих крыльев, на островах, где бушуют ветры, как, например, на островах Кергелен в южной части Индийского океана. На этих островах в процессе отбора преимущество получили насекомые с укороченными крыльями или вовсе без них по отношению к насекомым, имеющим нормально функционирующие крылья. Первым вряд ли грозит опасность быть унесенными ветром в открытое море, что означало бы их верную смерть. Этот пример показывает, что в определенных условиях окружающей среды воздействие естественного отбора направлено на потерю одной из структур. На подверженных сильным ветрам островах насекомые с остатками крыльев или вообще без них лучше приспособлены к условиям окружающей среды, чем насекомые с крыльями, но эта адаптация связана с потерей структур, то есть с дегенерацией. Главная проблема эволюционных исследований причинности - объяснение происхождения структур - в данном случае опять-таки не затрагивается.
Все описанное выше касается слепых и лишенных окраски пещерных животных. В этих случаях речь идет также о явлениях дегенерации, о так называемом "развитии по нисходящей".
Серповидноклеточная анемия
Серповидноклеточная анемия - это болезнь крови. Гомозиготные носители рецессивных генов серповидных клеток (ss) умирают в молодом возрасте. Продолжительность жизни гетерозиготных носителей генов (s+) относительно гомозиготных здоровых (++) лишь немного снижена. Так как в кровяных тельцах гетерозигот возбудители малярии все же не могут распространяться, комбинация аллелей s+ дает защиту от малярии и тем самым возможность выжить. Селекционное преимущество проявляется только в гетерозиготном состоянии, но и это лишь в чрезвычайных условиях окружающей среды (районы эпидемии малярии). Ген серповидной клетки не имеет шанса распространиться в популяции сверх определенной нормы (рис. 3.21).
Это было бы возможным только тогда, когда селекционное преимущество проявилось бы в гомозиготном состоянии, чего в данном случае не происходит. Поэтому этот пример для доказательства возможности развития от простого к сложному при помощи механизма отбора не подходит. Однако он является доказательством того, что селекционные преимущества отбора зависят от условий окружающей среды и могут привести к возникновению новых комбинаций аллелей в популяции.
Кроты
В популяции кротовых проводились измерения длины черепа до и после особенно холодной и долгой зимы. Было выявлено, что после длительного холода средняя длина черепа становится несколько меньше. Поскольку длина черепа может быть принята в качестве параметра измерения размеров тела животных, то этот показатель можно рассматривать как следствие неблагоприятного режима питания и отсюда - как показатель естественного отбора больших по размерам особей: маленькие особи обходились меньшим количеством пищи и поэтому при нехватке питательных веществ имели большие шансы на выживание. Естественно, должен существовать и какой-то минимальный размер животных.
Если даже изменение размеров тела связано с изменением состава аллелей в популяции, что, однако, в данном случае не доказано, то это говорит только о том, что, как и в случае с березовыми пяденицами, частота аллелей также меняется под влиянием изменившихся условий окружающей среды.
ДДТ- сопротивляемость насекомых
Последнее из наблюдаемых явлений естественного отбора, которое нам предстоит рассмотреть - это появление сопротивляемости ядам, используемым против вредных насекомых. Например, невосприимчивость мух яду ДДТ. При этом установлены явные различия генетического плана между мухами, имеющими иммунитет и не имеющими его. У резистентных штаммов выявлены следующие различия: пониженная проницаемость покровов и тканей для ДДТ, быстрое ферментативное разложение ДДТ в теле, способность откладывать больше ДДТ в измененном жире, пониженная восприимчивость к ДДТ нервной системы, изменение образа поведения, благодаря которому возможности контакта с ядом снижаются.
П.А.Тшуми (1975, с. 185) указывает на одну интересную точку зрения в рассуждении об этом селекционном явлении:
"Все исследования свидетельствуют о том, что интенсивная селекция изначально редких генетических вариантов изменила генетический состав популяций насекомых. Все резистентные (невосприимчивые к ДДT) мухи, иначе говоря, и есть потомки этих редких генотипов, которые избежали начавшихся массовых смертей. В случае отсутствия подобных редких вариантов сопротивляемость не проявляется. В действительности же имеются некоторые популяции мух, а также саранчи, которые до сих пор не развили никакого иммунитета".
Поэтому-то во всех известных случаях резистентные формы, уже имевшие ДДТ-иммунитет к началу отбора, никаких изменений не получили, за исключением частоты аллелей популяций, подвергнутых воздействию ДДТ.
Но это не является шагом к макроэволюционным изменениям. Хотя в некоторых случаях генетическая основа устойчивости неизвестна, однако вполне очевидно, что устойчивость, как правило, основывается на отсутствии генной функции. В качестве промежуточного вывода можно принять, что:
• изменение условий селекции может сопровождаться изменением частот аллелей в генофонде популяции. Однако это не равнозначно эволюционному шагу, соответствующему макроэволюционным изменениям.
Половой отбор
При помощи этого типа отбора пытаются объяснить появление у мужских особей вторичных половых признаков. Конкуренция за половое партнерство рассматривается как отдельный фактор внутривидового отбора. Еще Дарвин полагал, что особи с более выраженными половыми признаками скорее найдут себе партнера для спаривания и тем самым скорее смогут размножиться. Посредством такого полового отбора наряду с вторичными половыми признаками (например, оленьи рога, пышное оперение) должно возникнуть сложное поведение во время брачного периода. Такие признаки и поведение мужских особей привлекают внимание женских особей. Дарвин писал (1967/1859, с. 131): "Олень без рогов и петух без шпор имеют меньше шансов оставить многочисленное потомство".
Все вышеприведенные возражения против действенности эволюционных факторов отбора подходят и для полового отбора (изменения, обусловленные половым отбором, например, большая выраженность присутствовавших и ранее вторичных половых признаков, происходят только в рамках микроэволюционных изменений, поэтому нет смысла останавливаться на этом подробнее). Стоит лишь добавить, что ярко выраженные половые признаки могут и сократить шансы на выживание, так как их носители нередко лучше заметны и для своих врагов. Таким образом, половой отбор, происходящий внутри вида и межвидовая борьба могут прийти в противоречие друг с другом. Это обстоятельство приводит к дополнительным трудностям для понимания механизма возникновения вторичных половых признаков путем полового отбора.
Адаптагенез и анагенез.
Рассмотренные до этого места примеры создают достаточный эмпирический базис для второго вопроса, затронутого в этом разделе: в какой взаимосвязи находятся процесс повышения приспособленности и развитие от простого к сложному? Ответ на него состоит в том, что никакой взаимосвязи не наблюдается. Если естественный отбор обеспечивает лучшую адаптацию к условиям окружающей среды, то при изменении этих условий видоизменяются уже имеющиеся в наличии органы и признаки, причем иногда в сторону их развития, а иногда - дегенерации (движущий отбор). Теоретик и биолог Л. фон Берталанфи так поясняет этот аргумент:
"По моему мнению, нет ни искорки научного доказательства того, что эволюция в плане развития от простых организмов к сложным идет по пути возрастающей адаптации с селекционным преимуществом или с производством все большего потомства. Адаптация возможна на любом организационном уровне. Амеба, червь, насекомое или бесплацентное млекопитающее так же хорошо приспособлены, как и плацентарные, и если бы этого не было, то они бы уже давно вымерли" (Цит. по Kahle, 1984, с. 68).
Червь под действием меняющихся условий окружающей среды, становится посредством естественного отбора лучше приспособленным к новым условиям червем, но никогда - насекомым. Изменения в этом направлении не увеличивали бы его приспособленность в качестве червя, и по этой причине исключались бы естественным отбором (рис. 3.22).
О резистентности микроорганизмов к антибиотикам и других изменениях, обусловленных мутациями и отбором, будет сказано в следующем разделе (3.3.4).
Заключение
- Воздействие отбора является, как правило, стабилизирующим, но при изменившихся условиях окружающей среды оно - движущее и приводит (или может привести) к явному сдвигу частоты определенных аллелей, а также к дегенерации органон.
- Все результаты наблюдений относятся к области микроэволюции. Ни в одном из случаев не могло быть доказано возникновение качественно нового генетического материала.
- Уровень организации и степень адаптации живых существ не находятся ни в какой взаимосвязи с воздействием отбора, так как эту взаимосвязь невозможно доказать.
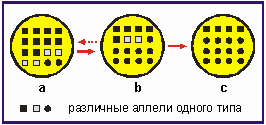
Рис. 3.15. Воздействие селекции на генофонд, а: состав аллелей определенного гена в определенной окружающей среде А: в: измененный состав аллелей после изменения условий окружающей среды (среда В) после смены нескольких поколении: с: дальнейшие изменения окружающей среды В в ходе смены поколений. Штриховая стрелка указывает на то, что изменения в генофонде под влиянием окружающей среды могут иметь и обратный ход, если соответственно изменятся и условия окружающей среды (опять среда А) и если не исчезнет ни одна аллель.
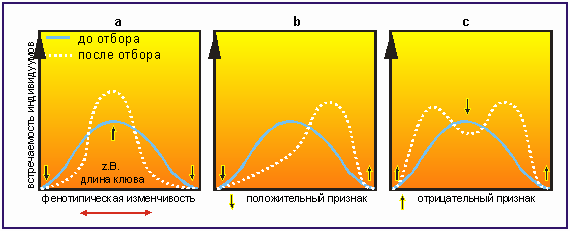
Рис. 3.16. a) стабилизирующий отбор; b) движущий отбор; с) дизруптивный отбор; по вертикали - частота встречаемости индивидуумов, по горизонтали - фенотипические вариации длины ключи; непрерывная синусоида - до селекции, прерывистая - после селекции. В качестве примера фенотипической вариативности можно выбрать длину клюва одного вида птиц. Стабилизирующий отбор воздействует так, что спектр изменения длины клюва в течение длительного времени держится в определенных узких рамках. Движущий отбор обуславливает смещение среднестатистической длины клюва, дизруптивный приводит к тому, что, скорее всего, выживут особи с самыми короткими и самыми длинными клювами.

Рис. 3.17. Светлые и меланиновые формы березовой пяденицы, (Рисунок THOMAS HILSBERG).

Рис.3.18 Подражательная форма у бабочки каллимы. Нижние части передних и задних крыльев являются подражанием форме листа дерева, хотя оба крыла возникают совершенно отдельно. (Вестфальский краеведческий музей, Мюнстер).

Рис. 3.19. Мимикрия безобидного шершневого бражника (внизу), который подражает шершню (вверху).

Рис. 3.20. Офрис пчелоносная.
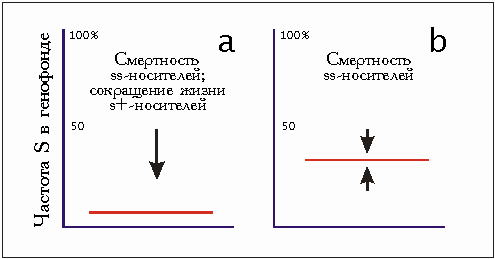
Рис. 3.21. Схема серповидноклеточной анемии, а - ситуация в районах, где отсутствует опасность малярийных заболеваний: аллель серповидных клеток s встречается редко; Ь - ситуация в районах малярийных эпидемий: аллель серповидных клеток встречается довольно часто (см. текст/. Но их частота не может превышать определенный порог (теоретически максимальный - 50 %). Аллель, тем самым, не может внедриться в генофонд популяции никогда (абсцисса безразмерна).
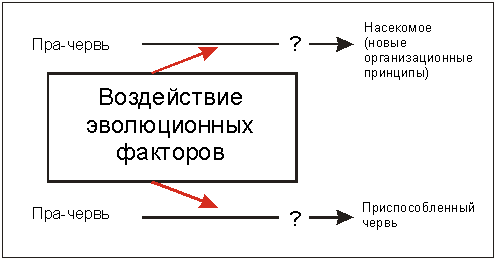
Рис. 3.22. Благодаря воздействию эволюционных факторов
согласно синтетической теории в течение длительных временных отрезков, например,
из гипотетического неспециализированного "прачервя" возникли насекомые. Эмпирические
данные указывают, однако, на то, что под действием эволюционных факторов из
неспециализированного "прачервя" может возникнуть только червь, специально приспособленный
к условиям окружающей среды.



