




История происхождения и развития жизни
4.3. Эффект мутаций и селекция на молекулярном уровне
Как уже было подробно описано, мутации должны поставлять "сырье" для эволюционного процесса. Посредством селекционных процессов всякий раз отбираются наиболее приспособленные к условиям окружающей среды индивидуумы. Не поддается оспариванию то, что селекция в природе является действенным фактором. Различие во мнениях состоит лишь в том, действительно ли могут описанные выше мутационные процессы произвести изменения в геномах и накопить в поколениях такие изменения, которые бы привели в дальнейшем к возникновению новых ферментных функций и, в конце концов, к более высокоразвитым формам. Чтобы прояснить эволюционно-теоретическую позицию, уместно привести здесь цитату из школьного учебника начального курса биологии (Knodrl и Bayrhliber, 1983, с. 403). В цитируемом отрывке дупликация генов описывается как механизм, посредством которого производятся (неактивные) копии генов (см. рис. 3.31).
"Многие из этих генов неактивны. В геномах человека и многих млекопитающих доказано наличие множества подобных неактивных генов, которые очень похожи на обычные структурные гены и предположительно от них и происходят. Неактивные, так называемые псевдогены, представляют собой некоторый "генетический запас" для будущей эволюции. Мутации в псевдогене не проявляются фенотипически. Со временем мутации накапливаются в псевдогене, и он изменяется. Если псевдоген становится в ходе эволюции активным и, тем самым, превращается в ген, то он может образовывать протеин с новыми функциями. Таким путем в процессе эволюции организмов возникает много новых свойств". На первый взгляд, эта модель выглядит довольно убедительно, однако часто бывает так, что отдельные подробности либо реальные примеры опровергают ее. Сила, а также слабость умозрительной модели становится ясной лишь тогда, когда она начинает касаться деталей проблемы. Это, естественно, касается в равной степени как моделей, ориентированных на эволюционную теорию, так и на модель сотворения. Чтобы обоснованно судить о процитированном выше высказывании, следует рассмотреть детально эту проблему в рамках эволюционно-теоретических предпосылок. Мы хотели бы рассмотреть это на примере из мира бактерий и из предполагаемого периода эволюции основополагающего процесса обмена веществ. Это нам представляется наиболее выгодным потому, что процессы обмена веществ и явления мутаций у бактерий исследованы в наибольшей степени (см. раздел 3.3,4). Их легко содержать в лаборатории, и они обладают настолько малым жизненным циклом, что представляют собой наилучший объект экспериментальных и теоретических исследований эволюционных процессов.
Получение бактериями энергии из "первичного бульона"
Обычно исходят из того, что являющиеся первыми живыми существами бактерии брожения берут свое происхождение из богатого органическими веществами "первичного бульона" (см. главу 5), где они получили необходимую для их выживания и размножения энергию при расщеплении "богатых энергией" органических молекул, таких, например, как сахар. Подобные бактерии существуют и сегодня. Принцип функционирования таких гипотетических "первозданных" бактерий изображен на рис. 4.9. Одноклеточная бактерия ограждена от "первичного бульона" мембраной из цитоплазмы. Бактерия питается сахаром, который попадает в клетку благодаря транспортировке его протеином. Это происходит не само собой, так как соответствующие сахара находятся в "бульоне" в крайне малой концентрации. Процесс усвоения сахара нуждается в энергии. О сегодняшних одноклеточных известно, что эта энергия поставляется, например, протоновым градиентом. Под градиентом протонов понимают неравную концентрацию протонов внутри клетки, по сравнению с внешней средой (в нашем случае - в "первичным бульоном"). В подобном неравновесии (разности потенциалов) черпается энергия. Наглядным примером могло бы служить неравновесие массы воды перед и за плотиной. Эта энергия может, например, использоваться для усвоения жизненно важного сахара - (1) на схеме 4.9А. Присоединение протонов посредством специального транспортного протеина начинает процесс усвоения сахара, подобно тому, как устремляющаяся из водохранилища гидростанции вода начинает вращение турбины. Внутри клетки сахар разлагается на более мелкие молекулы, к примеру, промежуточным продуктом может являться фумаровая кислота. В ходе подобных реакций расщепления (катаболических реакций) из АДФ образуются АТФ, а из HAД+ - НАДН + Н+. Растворимый фермент фумаратредуктаза (3) замещает фумаровую кислоту и НАДН + Н на янтарную кислоту, которая либо используется далее, либо в качестве побочного продукта покидает клетку (4). АТФ нередко называют энергетическим запасом клетки. Это богатое энергией соединение. Этой "валютой" клетка может "расплатиться" с требующим энергию синтезом (анаболические реакции). Без АТФ не может быть настоящего роста. И в ходе отбора лучше развиваются клетки, которые располагают лучшими механизмами образования АТФ. Многие клетки имеют в своей мембране протеин (5), который при расщеплении АТФ поставляет протоны внутрь клетки. С помощью АТФ-гидролазы выделяемая при расщеплении АТФ энергия "заряжает" вещества-переносчики, например, белки, сахара, которые переносят ее в те участки клетки, где она необходима. Мы уже знаем, для чего это полезно: жизненно важные сахара могут поглощаться, если во внешней среде больше протонов, чем внутри клетки, то есть если протоновый градиент находится за пределами мембраны клетки. Часть полученных от расщепления сахара молекул АТФ используется для производства протонового градиента. Функционирующая подобным образом клетка должна была, вероятно, появиться раньше всех имеющих к ней отношение эволюционных механизмов (хотя в эволюционном учении не имеется реалистичных предположений о том, как подобная клетка могла образоваться случайно, см. главу 5). И мы должны подробнее рассмотреть, как эта бактерия могла развиваться далее путем приобретения новых ферментных функций.
Новая метаболическая функция
Мы исходим из того, что новые функции только тогда образуются и утвердят себя. когда изменится окружающая среда и таким образом возникнет давление отбора (создадутся условия для начала селективного, в течение миллионов лет, процесса). Полагают, что на протяжении миллионов лет в процессе роста бактерий брожения наступило обеднение потребляемых ими из окружающей их среды органических молекул. Если попытаться изобразить этот "первичный бульон", то получится следующая картина: бактерии с течением времени "исчерпали" его и, должно быть, "страдают от голода". Тем самым можно объяснить преобладание сильного селективного воздействия, направленного на создание дополнительных механизмов для образования молекул АТФ. Наиболее удобной возможностью для этого было бы использование из "бульона" органических молекул, которые до сих пор не были еще расщеплены. Как это могло бы выглядеть, изображено на рис. 4.9В. Следует исходить из того, что в распоряжении могло быть ограниченное количество молекул сахара (1). Этот сахар был, как описано выше, расщеплен (2), но возникающая фумаровая кислота замещается янтарной не с помощью НАДН, а при помощи другой ферментной системы. Необходимые для этого протоны могут быть взяты из воды, но электроны берутся из некой новой ферментной системы (4). На внешней стороне мембраны муравьиная кислота расщепляется на СО2 протоны и электроны (4). Последние проникают сквозь мембраны клетки, передаются соединенной с мембраной редуктазой фумаровой кислоты и способствуют образованию янтарной кислоты. Важнейший результат этих реакций: на внешней стороне мембраны освобождаются протоны (4), внутри же клетки, напротив, протоны не освобождаются. В сумме это приводит к образованию протонового градиента: снаружи - много, а внутри - мало свободных протонов Мы помним, что в подобном протопопом градиенте черпается энергия, а источник энергии, созданный по такому принципу, с использованием такой схемы реакций, создается без расходования АТФ-молекул. Если же описанная выше АТФ-гидролаза направила бы свое действие в прямо противоположном направлении, то подобная бактерия располагала бы дополнительным источником энергии. Возникший в результате расщепления протоновый градиент используется таким образом для АТФ- синтеза. Можно с уверенностью сказать, что этот механизм задуман как переходная ступень для дальнейшего развития транспортной цепочки электронов. Эта ступень, бесспорно, имеет положительное селективное значение.
Необходимые "перестройки"
Опубликованные в международных научных журналах статьи о происхождении таких или подобных структур на этом месте заключают свои высказывания следующим: многими теоретиками эволюции проявление сильного давления отбора и приведенная выше попытка изобразить эволюционный процесс рассматриваются как вполне достаточное основание, на котором они строят принципиальную возможность подобной эволюции. Мы хотели бы, однако, продолжить рассуждения и спросить, какие, в частности, изменения в геномах бактерий должны произойти, чтобы возник изображенный выше вариант обмена веществ. Перестройку АТФ-гидролазы в АТФ-синтетазу можно было бы изобразить следующим образом: ген должен подвергнуться дупликации, а транскрипция дупликата должна быть предотвращена. Кроме того, давайте предположим, что два изменения цепочки аминокислоты сделают возможным синтез АТФ.
Современные АТФ-синтетазы являются многоферментными комплексами и состоят из много численных подразделений. Точный механизм АТФ-синтеза, приводимый в движение протона ми, до сих пор остается непонятным. Предположение о том, что посредством только двух точечных мутаций из некой достаточно просто устроенной АТФ-гидролазы, которая в современных организмах состоит из одного протеина, берет свое начало еще одна такая же "примитивная" АТФ-синтетаза, во всех случаях слишком далеко от биологической реальности. Впрочем, современные АТФ-гидролазы в существующих физиологических условиях необратимы.
Далее клетка нуждается в присоединенном к мембране протеине, который муравьиная кислота может расщепить и передать электроны сквозь мембрану на редуктазу фумаровой кислоты. Для этого должен быть расщеплен какой-нибудь предназначенный для этого "околомембранный протеин", а для этого, во-первых, должна произойти генная дупликация, во-вторых, необходимо предотвратить транскрипцию. Мутации, происходящие в покоящемся гене, должны вызвать ферментную активность в двух указанных направлениях. И, наконец, необходима, по крайней мере, одна соединенная с мембраной фумарат-редуктаза. Ген фумарат-редуктазы должен был опять подвергнуться дупликации с последующим предотвращением его транскрипции. Затем посредством мутаций он должен измениться так, чтобы стало возможным проникновение через мембрану клетки электронов, полученных в результате расщепления муравьиной кислоты, а не из НАДН. Сегодняшние фумарат-редуктазы из бактерий являются сложными агрегатами, состоящими из множества протеинов, которые кодируются не одним геном. Можно было бы привести еще множество условий, которые необходимо выполнит!,, чтобы придать достаточно реалистичности изображенному выше процессу функционального обмена, с тем чтобы можно было реально исходить из этой теории (например, образование механизма регуляции). Однако ради простоты изложения мы поступим следующим образом: допустим, что этих "перестроек" вполне достаточно. После того как произойдут необходимые мутации, удвоенные гены осуществляют транскрипции, и тем самым образуются протеины. Для этого также необходимы три мутации, которые снова активизируют соответствующие стартовые кодоны. В таблице 4.1 приводится количество необходимых мутаций. Для лучшего понимания следует помнить о том, что в ходе этого эволюционного процесса необходимо создать три каталитических центра. Сколько мутаций должно быть учтено при этом? МАНФРЕД АЙГЕН называет 15 изменений аминокислотных цепочек, которые необходимы для того, чтобы в протеине создать новый каталитический центр. Это можно было бы принять за реалистическую оценку данных, но мы бы хотели пойти в своих рассуждениях значительно дальше: только три аминокислоты должны быть изменены, чтобы произвести новую каталитическую единицу. Не должно, однако, возникать впечатления, что оценка идет лишь в пользу учения о сотворении. Итак, в общей сложности должно произойти, по крайней мере, 23 мутации, чтобы появились на свет желаемые мутанты.
Оценка вероятности
В самом начале следует еще немного упростить рассмотрение вопроса: мутации, подвергшиеся транскрипции, должны упраздниться, и к тому же все мутации должны произойти в период жизни бактерий одного поколения. Это требование является необходимым, так как только вся структура в целом действительно обладает селективным преимуществом. Легко можно предположить, что выпадение даже одной функции приведет к селективному ущербу для всей конструкции, и воздействие селекции пагубно отразится на подобном индивидууме. Без мутаций стартового кодона потребовалось бы только 17 независимых изменений ДНК. Так как мутации происходят случайно, и так как еще не было предложено в ходе поступательной эволюции ни одной способной функционировать промежуточной ступени, то можно считать, что явления независимы друг от друга, и тем самым умножить вероятность отдельных явлений. Важно ясно представлять себе, что в последующих рассуждениях и оценках следует учитывать аспект отбора. Частота вероятности всего явления зависит еще и от нормы мутаций. Предположим, что мутационная норма очень велика, около 10-5. Тогда вероятность всего процесса находилась бы при таких условиях в пределах 10-85. Это означает, что среди 1085 клеток в среднем можно ожидать лишь одну, которая имела бы желаемую комбинацию мутаций. Если бы все "первичные океаны" были бы полны бактерий, то через миллиард лет следует ожидать в общей сложности не более 1045 индивидуумов. Это означает, что желаемого события не следует ожидать, так как событие с вероятностью 10-85 • 1045 = 10 40 не только в математическом, но и в естественнонаучном понимании считается невозможным. Это число лежит далеко за пределами человеческих представлений, которые необходимы, чтобы сделать этот пример наглядным. Шанс случайно обнаружить булавку размером приблизительно 1 мм в объеме земли находится на уровне 10-32 ! Если читатель интересуется более подробным изложением предмета, то мы предлагаем ему обратиться к перечисленной в приложении литературе. Однако вернемся к началу наших рассуждений: если находящиеся в состоянии покоя гены в течение многих лет затем могли мутировать, то о какой одновременности событий может идти речь? Этой гипотезе предстоит борьба со следующими проблемами: как известно из экспериментов и концепции о динамике развития популяций, гены, не выполняющие никакой функции, находящиеся в стадии покоя, выбраковываются, особенно у бактерий, в исключительно короткий с геологической точки зрения срок. Уже одно это делает гипотезу несостоятельной, что, однако, для нас не должно иметь значения. Важнее то, что соответствующая мутация тоже не принесет организму никакого селективного преимущества, поэтому он, по сравнению с другими организмами, не будет быстрее размножаться (можно было бы даже предположить, что, испытывая большую "генетическую нагрузку", подобные организмы чуть ли не замедлят свое размножение). Так что "селекционный аргумент" " данном случае не подходит. Если общее число индивидуумов в некоем жизненном пространстве не увеличивается (а это в наших предпосылках очень важно), то частота появления индивидуумов, подвергшихся первой мутации, всегда будет одинаковой. Числовой пример пояснит этот' аргумент: предположим, что в иском общем первичном океане на некий заданный момент времени имеется 1035 индивидуумов. При все увеличивающейся в дальнейшем частоте мутаций равной 10-5, желательной мутации подвергнется 1030 индивидуумов. И доля их в последующих поколениях не изменится, разве только уменьшится. Даже горизонтальный генетический перенос ничего не изменил бы в этом положении, так как он не несет селективных преимуществ. Когда же, наконец, наступит вторая необходимая нам мутация, то она должна естественно произойти в той же самой бактерии Число индивидуумов, подвергшихся обеим мутациям, находится в пределах 1025. Третье изменение, независимо от времени своего наступления, затронет 1020 индивидуумов и пока произойдут еще 20 необходимых мутаций частота появления индивидуумов достигнет 10-65. К этому еще следует добавить и три находящихся в состоянии генетического покоя гена, которые должны быть одновременно активизированы, а это еще уменьшит общую вероятность до 1080. Даже если принять во внимание, что мы неточно предположили время эволюции, это не уменьшит достоверности этих результатов Постулат о находящемся в состоянии покоя гене и о сдвиге времени наступления мутаций сводится, в конце концов, к уменьшению вероятностей что можно объяснить дополнительными необходимыми мутациями стартового кодона.
Все приведенные выше высказывания изложены кратко а заинтересованному читателю предлагаем для дополнительного углубленного изучения работы, указанные в перечне литературы
Возможные возражения
Теперь следует подробнее коснуться некоторых возражений против изложенных в этом разделе концепций (в частности оценки вероятности) Результат, очевидно, зависит в решающей степени от нормы мутаций. Реально ли намечать более высокую норму мутаций? Известно, что клетка посредством сложного механизма восстановления сдерживает частоту мутаций на низком уровне. Частота в 10-4 или даже 10-3 привела бы в короткий срок к вымиранию этой линии клетки, так как наслоение в большинстве своем негативных мутации привело бы к так называемой "катастрофе ошибок". Это могло бы произойти и после более длительного периода времени при общей норме мутаций 10-5 . Так как исходят из тою, что существовавший к тому времени генетический аппарат в общих чертах был похож на современный, то дальнейшее повышение нормы мутаций можно исключить. Далее, норма мутаций зависит еще и от величины подвергшеюся мутации гена - чем длиннее ген, тем больше шанс замещения любого основания гена. Но, во-первых это не изменит величины больше, чем на каких-нибудь два-три порядка, во-вторых, невозможно допустить что бы эти 'примитивные" гены были, в среднем, очень длинными. И, наконец, более высокая общая вероятность изменения гена не означает, что произойдут именно необходимые изменения и именно на предусмотренном нуклеотиде, а не где-либо еще в гене.
Здесь ясно видна граница современного уровня знаний. В приведенной выше оценке мы исходили из гена в общем виде или из протеина. Точнее было бы связать эту оценку с нуклеотидами, чьи мутации и производят желаемые изменения, и лишь потом рассчитывать частотность мутации. К сожалению, таковые неизвестны, поэтому придется довольствоваться приведенной выше оценкой. Поскольку эта оценка построена на замерах мутационных явлений в бактериях, она дает достаточно реалистичный результат, хотя в ней не упоминается отдельно нуклеотидный обмен.
В качестве вспомогательного средства для возникновения многофакторных мутации часто называют горизонтальный генный перенос. Разве не могло быть так, что в одной бактерии мутации подвергся некий ген, во второй бактерии - другой ген, и что благодаря горизонтальному генному переносу оба гена соединяются и находят свое выражение? Эта гипотеза, во-первых, не учитывает, что частота для изменений даже одного гена чрезвычайно мала. Во-вторых, следует задаться вопросом, насколько велики шансы того, что в первичном океане как раз оба эти индивидуума, в которых случайно развились подходящие друг другу псевдогены, встретятся и вследствие этого произойдет генный трансферт (что относится к довольно редким явлениям)? Однако это не какой-то единичный генный трансферт: подвергаться переносу должны около сотни генов "примитивной бактерии". Но и это останется безуспешным если гены не разместятся на нужном месте в геноме. Не вдаваясь далее в детали, можно понять, что горизонтальный генный перенос не может способствовать развитию от низших форм к высокоразвитым. С другой стороны, посредством этого может быть достигнуто быстрое и эффективное приспособление (см раздел 3.3.4.)
Иногда встречаются предположения о том что в паре с "молчащей" мутацией могла выступить случайно позитивная мутация которая обеспечивает индивидууму селективное преимущество, но это не должно иметь никакой взаимосвязи с рассматриваемым нами эволюционным процессом. Наш псевдоген в известной мере принимается в расчеты по системе "Huckepack" (система снятия кандидатуры на выборах в пользу партнера по коалиции), что предполагает сверхпропорциональный рост его частоты в популяции. Тем самым создается гораздо более высокая вероятность его дальнейшей мутации. Но, в общем, эта выдуманная модель не дает большей вероятности, так как позитивная мутация, к которой должен присоединиться псевдоген, проявляется даже при малой вероятности, способной соответственно ухудшить общий результат. И, наконец, следует обсудить возможность того, что мутация может произойти в уже имеющемся генетическом "фрагменте" и, тем самым, заменить две или три отдельных мутации. Это теоретически возможно в экзон/интрон-структуре генома (см. рис. 4.4). Но поскольку мы рассмотрели пример из мира бактерий, то этот аргумент отпадает сам собой, так как в генах бактерий не выявлено экзон/интрон-структуры. Сможет ли этот путь обеспечить клеточным живым существам (содержащим ядра в клетках) развитие к более высоким формам, мы обсудим в разделе 4.4.
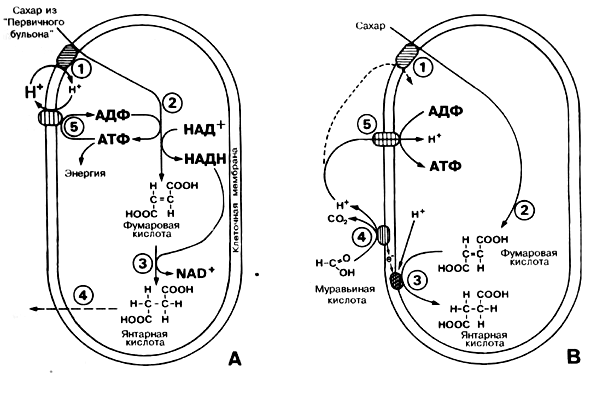
Рис 4.9. А: Основной путь добычи энергии бактериями брожения из "первичного бульона покачан и левой части схемы. (1) - сахар с помощью градиента протонов транспортируется через мембрану и расщепляется в процессе гликолиза (2, 3). (4) - конечный продукт покидает клетку. Возникающая при гликолизе АТФ частично (5) используется для того, чтобы восстановить по ту сторону цитоплазматической мембраны протоны, которые опять понадобятся для усвоения следующей порции сахара. Как такая бактерия могла возникнуть случайно - неизвестно. В: В качестве шага на пути эволюции можно было бы предположить расщепление муравьиной кислоты на внешней стороне цитоплазматической мембраны. Через мембрану передвигаются электроны, и непрямым путем может возникнуть градиент протонов, который клетка сможет использовать как для транспортировки сахара, так и для ЛТФ-синтеза. (I) - поглощение сахара, (2) - гликолиз, (3) - промежуточные продукты гликолиза редуцируются (восстанавливаются) посредством электронов (4), происходящих из муравьиной кислоты, и конечный продукт опять покидает клетку. (5) - образование АТФ посредством присоединения прото-
|
Число необходимых мутаций |
|
|
Образование АТФ-синтетазы: |
|
|
Дупликация гена |
1 |
|
Инактивация стартового колона |
1 |
|
Изменение двух аминокислот |
2 |
|
Образование дегидрогеназы муравьиной кислоты: |
|
|
Дупликация гена |
1 |
|
Инактивация стартового кодона |
1 |
|
Образование активной части фермента для расщепления муравьиной кислоты |
3 |
|
Образование активной части для того, чтобы можно было редуцировать фумаровую кислоту |
3 |
|
Преобразование редуктазы фумаровой кислоты: |
|
|
Дупликация гена |
1 |
|
Инактивация стартового кодона |
1 |
|
Образование протеина мембраны |
3 |
|
Образование активной части, которая может принимать электроны из муравьиной кислоты |
3 |
|
Активация стартовых сигналов для транскрипции |
3 |
|
Итого: необходимых мутаций |
23 |
Таблица 4.1. Мутации, необходимые для того,
чтобы из бактерии брожения (Рис.5.9) развилась первая клетка, которая приобретает
энергию за счёт градиента протонов. Пояснения в тексте.



