




История происхождения и развития жизни
6.4. Гипотеза об эндосимбионтах
В целом организмы можно разделить на две группы: на организмы, клетки которых содержат настоящие клеточные ядра, и организмы, которые этим свойством не обладают. Первые называются эукариотами, вторые - прокариотами. К прокариотам относятся бактерии и сине-зеленые водоросли. Эукариоты объединяют все остальные одно- и многоклеточные живые существа. В противоположность прокариотам, кроме обладания клеточными ядрами, эти существа отличаются выраженной способностью к образованию органоидов. Органоиды - это разделенные мембранами составные части клеток. Так, самыми большими клеточными органоидами (по крайней мере, различимыми в световой микроскоп), которыми обладают эукариоты, являются митохондрии, а растительные организмы обладают еще и пластидами. Митохондрии и пластиды большей частью отделены от цитоплазмы клетки двумя мембранами. (Некоторые подробности строения см. на рис. 6.13). Митохондрии часто называют "силовыми станциями" эукариотических клеток, так как они играют
большую роль в образовании и превращении энергии в клетке. Пластиды для растений не менее важны: хлоропласта, которые являют собой основной тип пластид, заключают в себе механизм фотосинтеза, который осуществляет превращение солнечного света в химическую энергию.
Так как прокариоты устроены значительно проще, чем эукариоты, то по канонам эволюционной модели считается, что прокариотические живые существа возникли раньше. Этим объясняется употребление приставки "про" (в смысле "до того"). Более нейтральным названием было бы, вероятно, "акариоты" ("а" = "не"). В ходе дальнейшей эволюции одноклеточные живые существа должны были, вероятно, когда-то совершить переход от про- к эукариотам. Один из важных частичных аспектов этого шага пытаются объяснить так называемой эндосимбиотической гипотезой (ЭСТ). В своем первоначальном виде она была выдвинута еще в 1883 году шимпером. Она являлась не единственной попыткой объяснения, но считалась на тот момент наиболее вероятной. Согласно этой теории, митохондрии происходят от аэробных (дышащих кислородом) бактерий, а хлоропласты - от фотосинтезирующих сине-зеленых водорослей, которые внедрились в "хозяйскую клетку" (в предка-прокариота) и в ходе эволюции превратились там из симбионтов (эндосимбионтов, точнее цитосимбионтов = клеточных симбионтов) в клеточные органоиды. Некоторые биологи предполагают (см. рис. 6.14), что жгутиково-центриольная система эукариотов произошла от ранее самостоятельных прокариотов (типа спирохет). Следующие факты рассматриваются в качестве наиболее важных для поддержки ЭСГ:
- Митохондрии и пластиды происходят путем деления им подобных. Клетка не может образовать вновь эти органы, если они утеряны.
- Обладание двойной мембраной создает впечатление, что речь идет о "внедренной клетке", чья мембрана при внедрении оказалась окруженной мембраной клетки-хозяина.
- Внутренняя мембрана митохондрии содержит липид кардиолипин, который, кроме этого, встречается только в мембранах прокариотов. Внешняя мембрана, напротив, как другие мембраны эуцитов (так называются клетки эукариотов), содержит холестерин, которого нет ни во внутренних мембранах, ни у бактерий.
- Митохондрии и пластиды содержат ДНК, которые, как и у прокариотов, "голые", что означает "не соединенные с протеинами", и часто имеют кольцеобразную форму. Они обладают также своим собственным механизмом синтеза протеинов, составные части которого (рибосомы, т-РНК и РНК-полимеразы) соответствуют составным частям прокариотов.
- Рибосомная РНК пластид или, соответственно, митохондрий имеет большое сходство с РНК прокариотических рибосом.
- Митохондрии реагируют на некоторые (не на все) направленные против бактерий антибиотики.
- Среди существующих ныне организмов встречаются случаи симбиоза между одноклеточными жгутиковыми, не имеющими пластид, и клетками водорослей, которые могли бы служить моделью определенной ступени филогенетического процесса эндосимбиоза.
Амеба Pelomyxa palustris не имеет митохондрий, но обладает эндосимбионтными бактериями. В некоторых школьных учебниках она представлена в виде промежуточного звена, которое призвано сделать наглядным превращение бактерий в митохондрии. Однако Pelomyxa для этой цели не подходит, так как её бактерии не поставляют ей АТФ, а лишь освобождают молочную кислоту от ядовитого продукта обмена веществ. Эндосимбионты не занимаются ни клеточным, ни бактериальным дыханием и не выполняют функций митохондрий.
Из приведенного выше перечня становится ясно, что основным доказательством в пользу этого предполагаемого эволюционного процесса являются пункты, в которых говорится о "сходстве" (пункты с 1 по 6. см. также 6.1). Наличие симбиоза (пункт 7 в приведенном выше перечне) не доказывает наличие перехода, но иллюстрирует лишь первую ступень, которая при попытке зволюционно-теоретического толкования несет с собой новые проблемы.
Сходству, однако, противостоят глубокие различия (мы ограничимся сравнением бактерии - митохондрии):
- бактерии выделяют протеины, митохондрии - нет;
- митохондрии принимают протеины и ЛДФ извне, бактерии - нет;
- определение генетического кода ДНК митохондрий отличается некоторыми кодонами (у человека - четырьмя) от ДНК бактерий (и даже не у всех до настоящего времени исследованных организмов они одинаковы). Кроме того, этот гипотетический переход несет с собой целый ряд сложностей:
- отличающийся от "универсального кода" кодон влечет за собой все возрастающее число происходящих параллельно мутаций в рибосомах, в некоторых т-РНК и т-РНК-синтетазах и в ДНК митохондрий, в противном случае, вся система не могла бы функционировать;
- большая часть протеинов митохондрий кодируется геномом, то есть, ДНК, содержащейся в ядро. Поэтому в рамках этой теории существует предположение, что с течением времени гены постепенно переселяются из митохондрий в геном ядра (соответственно, это касается и пластид). Всякое отдельно взятое "переселение" влечет за собой много происходящих независимо друг от друга, "сочетающихся друг с другом" мутаций. В этих случаях, поскольку селекция лишь после огромного числа независимых мутаций может оценить "результат" как позитивный и тем самым "зафиксировать" его становится возможной оценка вероятности перехода от одной ступени к следующей (селективно-положительной). По этим оценкам степень вероятности ничтожно мала. Это означает, что, согласно современному уровню науки, следует рассматривать эволюционное происхождение эукариотов в рамках названной выше гипотезы как в высшей степени невероятное.
Так как эта теория считается относительно остальных наиболее вероятной, то будем считать, что мы включили в нее и оценку конкурирующих теорий об эволюционном происхождении эукариотов. Наиболее заинтересованным читателям предлагаем обратиться к списку дополнительной литературы (schneidfr, 1985). О том, как производится подобная оценка вероятности, описано в главе 4.
Упомянутое сходство между митохондриями (или, соответственно, пластидами) и бактериями (или сине-зелеными водорослями) может и должно быть связано с функциональной необходимостью. Задача дальнейших исследований - выявить, какой смысл имеет то, что генетический код митохондрий частично иной, чем код клеточного ядра, и поэтому аппарат для синтеза протеина больше похож на аппарат бактерий или, соответственно, сине-зеленых водорослей, чем аппарат цитоплазмы. Посмотрим, может ли учение о сотворении дать ответ на этот вопрос.
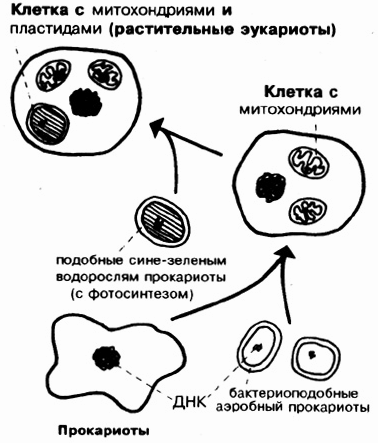
Pис. 6.14. Схема эндосимбиотической гипотезы, сильно упрощенная.(По
Mohn, 1934, u Margulis.)



