




История происхождения и развития жизни
3.5.2. Теория дополнительного типогенеза.
Теория дополнительного (аддитивного) типогенеза является еще одной попыткой разрешить проблему макроэволюции. Под аддитивным типогенезом понимают следующие гипотезы: новые организационные формы возникают не в ходе одновременного изменения затронутых организационных признаков, а путем последовательно происходящей замены отдельных признаков. И только по прошествии длительного периода времени вследствие суммирования отдельных изменений рождается новый организационный тип (рис. 3.63).
Согласно этим представлениям, в ходе развития млекопитающих из рептилий различные типичные признаки млекопитающих, такие как волосяной покров, молочные железы, три слуховые косточки, способ передвижения и т. д., развивались постепенно, друг за другом. Именно процесс развития млекопитающих из рептилий мог бы быть доказан с помощью ископаемых. Так, например, известны такие ископаемые рептилии из группы терапсид, которые имеют два челюстных сустава: первичный - рептилий, вторичный - млекопитающих (см. раздел 3.3.6) Среди терапсид имеется много видов рептилии с целым рядом признаков млекопитающих (насколько это вообще можно установить по окаменелостям). Еще одним указанием правильности этого положения эволюционной теории считается существование переходных форм (мозаичных типов). Подобные переходные формы соединяют в себе признаки обеих больших систематических групп (см. раздел 8.7). Но это никоим образом не доказывает, что в данном случае имеет место родовая взаимосвязь, что они связаны между собой генетически. Прежде всего, процесс образования отдельных организационных признаков даже с помощью теории аддитивного типогенеза остается неясным, что и демонстрирует пример возникновения слуховых косточек у млекопитающих (см. раздел 3.3.6). Теория аддитивного типогенеза разделяет проблему образования типов на множество отдельных вопросов о возникновении в процессе эволюции отдельных новых структур, т.е. просто приводит снова к проблеме макроэволюции.
Даже ссылка на коренное изменение условий окружающей среды или на освоение новых экологические зон не поможет, так же как и предположение, что макроэволюционные процессы происходили в относительно малых популяциях, что и могло вызвать генный сдвиг. Ни подобные предположения, ни теоретические вспомогательные конструкции не внесли прояснения в проблему эволюционных механизмов, которой как раз и занимается исследование причинности в эволюции.
В этой связи представляет интерес еще один часто цитируемый
пример: возникновение хрусталикового глаза. В рамках эволюционной теории предпринимаются
попытки объяснить эволюцию этого сложного органа восхождением по функциональным
ступеням: в качестве исходного пункта развития принимается пучок зрительных
нервов, и дальнейшее развитие прошло затем через стадии плоского глаза, чашеобразного
глаза и заглубленного глаза к хрусталиковому. Существование этих организационных
ступеней является, конечно, необходимым, но никак не достаточным условием для
постепенного развития. Перечисление возможных промежуточных ступеней, однако,
не объясняет ни переходов от ступени к ступени, ни лежащих в их основе генетических
изменений.
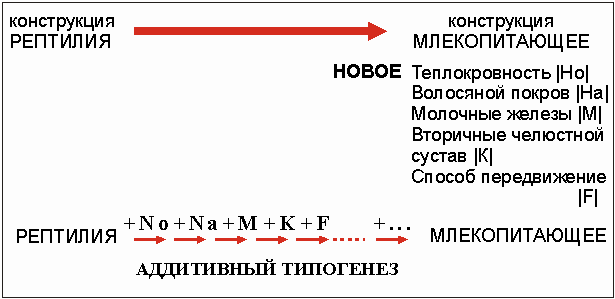
Рис. 3.63. Аддитивный типогенез. R рамках эволюционной теории
переходы между различными типами конструкции представляются следующим образом:
отдельные новые структуры появляются одна за другой, то есть последовательно.
Но н примере перехода от рептилии к млекопитающим выбранная последовательность
суммирования новых структур составлена абсолютно произвольно!



