




История происхождения и развития жизни
9.3. Австралопитеки
Австралопитеки - промежуточное звено между животным и человеком. Между рамапитеками и различными формами австралопитеков до настоящего времени наблюдался разрыв в цепи ископаемых находок более, чем в 4 миллиона лет (рис. 9.1). Многие палеонтологи надеются с помощью находок новых окаменелостей из слоев, относящихся к временному периоду между 8 и 4 миллионами лет, получить новый импульс в процессе познания, чтобы осмыслить это непонятное "становление человека". Австралопитеков относят к семейству человекообразных, считают прямым родственником человека. В этом разделе мы разберем, рассматривая австралопитеков в качестве высокоспециализированной группы приматов, почему их особенности нельзя интерпретировать как признаки процесса антропогенеза.
Исторический аспект проблемы изучения австралопитеков
Современную дискуссию об австралопитеках невозможно понять, не осмыслив исторического фона, на котором происходили исследования. Поэтому углубимся в историю их открытия. К началу нашего столетия среди специалистов господствовало мнение, что линия развития человека должна быть очень длительной. Базируясь на чисто теоретических предположениях, ученые считали, что сначала развился мозг человека, а потом вертикальная походка. В большей степени этот вывод был обязан нераспознанной в течение 40 лет фальсификации черепа древнего человека, составленного из верхней части человеческого черепа и нижней челюсти обезьяны. Открытый в 1925 году вид Australopithecus africanus был определен как предок шимпанзе. Лишь с 1950 года начало пробивать себе дорогу мнение о том, что сначала произошла эволюция вертикальной походки и лишь потом - эволюция человеческого мозга. Тем самым австралопитеки с их относительно малым размером черепа и некоторыми признаками, свидетельствующими о вертикальной походке, стали наиболее вероятными кандидатами на роль предков человека. Имеются и другие исследователи (среди них Лики), которые не считают вид Australopithecus предком человека и считают, что человек произошел от более древнего независимого рода (рис. 9.8C)
Основная трудность палеоантропологии состоит в нехватка ископаемого материала. !)то приводит к тому, что некоторые гипотезы остаются на чисто умозрительном уровне. И весьма вероятно, что на основании полученных впоследствии данных, они подвергнутся изменениям. Наглядным примером этому является так называемый "человек из Небраски", облик которого в 1922 году реконструировали на основании найденного ископаемого зуба. Пятью годами позже пришлось признать, что этот зуб принадлежал вымершей свинье. Точно также потребовалась ревизия толкованию отпечатков ног, оставленных в слоях меловой формации в местности Пейлукси-ривер, штат Техас. Эти отпечатки обнаружены в непосредственной близости со следами, оставленными ящерами. Многие сторонники модели сотворения считают их отпечатками следов человека. Однако, по мере их выветривания становится ясно, что они принадлежали не человеку, а, предположительно, до сих пор неизвестному виду трехпалых ящеров. К сожалению, в области палеоантропологии нередко встречается феномен "научного обмана". Подобный обман в определенных условиях может оставаться нераскрытым в течение десятилетий.
Описанные в приведенных выше примерах случаи оказали достаточно сильное влияние на развитие соответствующих гипотез, однако прогресс в исследованиях вносил в них все новые и новые коррективы. Было бы несправедливым на основании подобных, достаточно редких случаев обвинить всю палеоантропологию в сознательной фальсификации или злонамеренных ложных трактовках. Во всем этом проявляется то, что рассуждения в области всех теорий о происхождении самым тесным образом связаны с интуицией и воображением исследователей. Поэтому следует проявлять здоровую долю осторожности, прежде чем ту или иную трактовку признать "истинной".
Гипотезы о таксономическом и филогенетическом положении австралопитеков.
Преодолев всю неразбериху, которая царила на протяжении всей истории с определением положения австралопитеков на родословном дереве, большинство ученых на сегодняшний день считают их прямыми предками человека. Однако, до сих пор не существует единства мнений о том, какой именно вид австралопитека является предшественником человека. Как правило, в качестве предков человека рассматривают австралопитеков "изящных". Однако встречаются исследователи, которые приписывают эту роль австралопитеку "коренастому" - robustus. К виду австралопитеков "изящных" относят Australopithecus afarensis и Australopithecus africanus. И на сегодняшний день наиболее распространенным и популярным является мнение о том, что именно A. afarensis состоит в непосредственном родстве с человеком по восходящей линии (см. рис. 9.8А). Поэтому сначала обсудим комплекс ископаемых находок, относящийся к виду A. afarensis, к которому среди прочих причисляют на удивление полно собранный скелет, известный под именем "Люси".
Australopithecus afarensis
Палеоантрополог джохангон нашел в местности Хадар (Эфиопия), кроме "Люси", останки еще 13 человекообразных. В одной из первых публикаций на эту тему ученый излагал мнение о том, что эти кости принадлежат трем различным видам (Homo. A. africanus и Д. robustus), но вскоре пересмотрел свое мнение и пришел к выводу, что речь в данном случае идет об останках одного вида с сильно выраженным половым диморфизмом (мужские и женские особи имели значительные различия в морфологии, подобные или даже более выраженные, чем наблюдаемые у горилл, см. рис. 6.19 а, б). Он объединил свой материал с окаменелостями, обнаруженными в местности Лаетоли, расположенной в 1000 км от Хадара. Возраст этих окаменелостей считался на 0.75 миллиона лет большим, чем возраст ископаемого материала из Хадара. Именно этот вид (afarensis) он определил в качестве непосредственного предка Homo вместо ранее предложенного Л. africanns. джохангон приписывает этому виду atarensis наряду с примитивным обезьяноподобным строением черепа, способность передвигаться вертикально, и полагает при этом, что нашел вполне подходящее промежуточное звено в развитии человека из обезьяны.
Между тем число критиков, атакующих точку зрения джохангона, со всех сторон растет. Многие из них вообще отказываются трактовать находки джохангона как отдельный новый вид: одна часть ученых склонна видеть в них представителей уже известного вида Д. africanus, другие же предполагают, что ископаемый материал "afarensis" следует отнести к двум (A. africanus и A. robustus) или даже более видам (в том числе и к Homo), как вначале считал и сам джохансон. Последняя попытка соответствует представленному на рис. 9.8 варианту D. Резкой критике подверглось также утверждение джохангона о том, что Д. afraensis имели вертикальную походку. Имеется множество доказательств (кости плеча, форма грудной клетки, очертания подвздошной кости, тазобедренный, коленный и голеностопный суставы) того, что "Люси" проявлял и явно выраженную приспособленность к жизни на деревьях и, возможно, имел способ передвижения, похожий на способ передвижения шимпанзе. Обнаруженные в Лаетоле следы стоп одни считают принадлежащими человеку, другие же, напротив, видят в них отпечатки следов лазающих форм.
Ряд ученых, обычно из числа критиков джоханcона, видят в классических формах Д. africanus (рис. 9.9) наиболее подходящих кандидатов в прямые* предшественники человека. Д. africanus должен был произойти от A. afarensis, либо быть с ним одним и тем же видом (рис. 9.8 В). Но и это мнение оспаривается. Сторонники родословного дерева по варианту А на рис. 9.8 указывают на то, что Д. africanus обнаруживает известный уровень специализации, который объединяет его скорее с A. robutus, чем с Homo. Наряду с этим, окгнард (1984), который не является сторонником ни варианта Л, ни варианта В (рис. 9.8) родословного дерева, пытается доказать с помощью многовариантного анализа признаков (математический метод суммирования и классификации различных признаков), что австралопитеков можно выделить из всех других родов в отдельную специфическую группу. Опираясь на это, он приходит к выводу, что их вообще нельзя рассматривать как предшественников человека.
Двигательный аппарат, по мнению Оке Нарда (1984), указывает на весьма специфический способ передвижения австралопитеков, сравнимый разве что со способом передвижения орангутана (лазанием по ветвям деревьев на руках без помощи нижних конечностей). В тазобедренной области наблюдается своеобразная комбинация признаков, которая, с одной стороны, позволяла вертикальную походку, а с другой - демонстрировала приспособленность этих особей к карабканью и лазанью при помощи передних конечностей но ветвям деревьев. В последнее время ученые с разных сторон обращают внимание на то, что австралопитеки не только передвигались по земле на двух ногах, но также могли обитать и на деревьях.
Утверждение о том, что Д. africanus является предком человека, подкреплялось еще и тем, что в течение длительного времени считали, будто он производил орудия производства. Однако ни в одном случае установить убедительную связь между австралопитеками и орудиями производства не смогли, так как в тех же самых или в одинаковых с ними слоях обнаруживали непременно останки человека. Поэтому этот аргумент из научных дискуссий был повсеместно исключен. Остальные ученые, которые не рассматривают на сегодняшний день представителей австралопитеков в качестве предшественников человека, также считают, что линия развития вида Homo берет свое начало гораздо раньше, чем это показано на рис. 9.8 в вариантах А и В. Они строят свои воззрения, опираясь на ископаемые экземпляры, якобы насчитывающие 5 миллионов лет, которые можно отнести к роду Homo. Сторонником более древней линии развития человека выступил среди прочих и Лики, чья схема родословного дерева изображена в варианте С (рис. 9.8).
Местоположение различных видов австралопитеков на родословном дереве относительно друг друга.
Вплоть до лета 1986 года считалось, что эволюция массивных австралопитеков (к ним относятся A. robustus и A. boisei) идет от представителей грациальных австралопитеков, ибо эта ветвь в эволюции человекообразных сохранилась наилучшим образом. Эта эволюционная линия развития, казавшаяся до сих пор весьма убедительной, была поставлена под сомнение с того момента, когда в Кении был обнаружен череп, датируемый 2,5 миллионами лет. На удивление хорошо сохранившийся череп имел большое сходство с гиперробустным Л. boisei, однако появился он "слишком рано", приблизительно на 1 млн. лет раньше, чем это допускают рамки общедоступных представлений о родословном дереве. Этот череп, отмеченный на рис. 9.8 A - D звездочкой, невозможно без должного осмысления вмонтировать в привычные схемы родословного дерева, так как он является убедительным свидетельством того, что A. boisei не мог произойти от A. robustus. как это предполагалось ранее. Эта находка привела к тому, что все ученые вынуждены были в той или иной степени видоизменить варианты их родословных деревьев. Суть изменений сводилась к следующему: эволюцию человекообразных следует увеличить на больший временной период, нежели предполагалось ранее, и допустить наличие не менее трех параллельных ветвей развития - двух линий австралопитеков и одной человеческой. Те ученые, которые в ископаемом материале вида afarensis наряду с gracilis-формами признавали и наличие rоbustus-форм, попытались сконструировать схему, похожую на вариант Д, в которую без проблем можно было бы включить и последнюю ископаемую находку из Кении. Несмотря на то, что эта находка прямого влияния на теорию эволюции человека не оказала, она все же ввела теоретиков в затруднение: признав трехлинейность развития, следует согласиться, что некоторые признаки должны были возникнуть троекратно и независимо друг от друга. Подвергнутое изменениям родословное дерево стало похоже на "куст форм", что в большей степени соответствует модели сотворения.
Мозг австралопитеков.
Высокоразвитый мозг человека существенно отличается по ряду признаков от мозга человекообразных обезьян. Австралопитеки соединяют в себе некоторые признаки обеих групп настолько специфическим образом, что эту комбинацию можно рассматривать и как промежуточную форму, и как отдельную, не имеющую аналогов в современном мире. На рубеже прошлого столетия ученые придавали для филогенетической классификации абсолютной величине мозга еще большее значение, чем в настоящее время, так как все остальные комплексы признаков они соотносили с этим параметром. Для определения объема мозга у ископаемого экземпляра применяются различные методики, имеющие целью получить как можно более близкое к действительности значение. И все-таки, поскольку на определение объема мозга большое отрицательное влияние оказывает недостаточное количество ископаемого материала, всевозможные деформации или неточности в реконструкции, связанные с техническим несовершенством, мы вынуждены пользоваться в большей или меньшей степени приблизительными данными. В случае с австралопитеками можно исходить из того, что объем их мозга в зависимости от величины тела колебался между 440 и 530 см2. Но эти же значения входят и в диапазон колебаний величины мозга африканских больших обезьян (см. таблицу 9.1). Значительный объем мозга не обязательно свидетельствует об интеллекте. Решение вопроса об интеллекте предусматривает корреляцию объема мозга с общим весом тела. А определение веса тела ископаемых форм является делом еще более сложным, чем определение объема мозга. Некоторые трудности привносит и понятие "относительной величины мозга". Из этого соотношения невозможно вывести степень организации или степень развития. Отношение веса тела к объему мозга, например, у обезьян-капуцинов явно выше, чем у человека (таблица 9.1).
Причина ошибочных выводов кроется в том, что остается неучтенным влияние величины тела на величину мозга. Так, с увеличением размеров тела изменяются и пропорции черепа. Таким образом, соотношения различных частей тела меняются не изометрически (пропорционально), а аллометрически. До настоящего времени не было предложено ни одной величины для определения уровня организации мозга и его развития.
Указывает ли изменение пропорций на принципиально иное строение, например, на другую организационную ступень, или это простое увеличение размеров - ответы на эти вопросы можно получить, только лишь сравнивая аллометрические графики родственных видов. Промеры и расчеты Xеммера (1967) показали, что все черепа австралопитеков и человека (включая Homo erectus) располагаются на двух абсолютно разных аллометрических Прямых, которые параллельны друг другу. На основании этого Хеммер делает вывод, что для перехода от австралопитека к человеку необходим транспозиционный скачок (см. тему "Макроэволюция" раздел 5.3). Различный объем черепа внутри одной аллометрической группы не является для австралопитеков показателем прогрессирующего процесса "гоминизации". Даже объем черепа вида Australopithecus habilis теряет свое кажущееся "промежуточное положение". Выводы Хеммера противоречат бытующему представлению о том, что емкость черепной коробки на стадии австралопитеков постепенно увеличивалась.
В хорошо сохранившихся экземплярах черепов по внутренним отпечаткам можно судить о наличии, величине и местоположении определенных структур мозга. И в этой области ученые проводили всевозможные исследования. Интерпретации отдельных структур мозга, предложенные ими, были весьма спорными, порой взаимоисключающими. Одни усматривали в отпечатках на внутренней поверхности черепа признаки человеческой организации мозга, другие подчеркивали сходство с современными большими обезьянами и отсутствие доказательств способности к речи (критический обзор, врандт, 1982).
В последних работах на эту тему, основываясь на изучении сохранившихся отпечатков мозга африканских человекообразных, ученые делают вывод о том, что австралопитеки по своей морфологии были похожи скорее на обезьян, так же как и некоторые представители habilis - комплекса (см. раздел 9.4), в то время как Homo 1470 проявляет абсолютно человеческий образ поведения (см. рис. 9.10).
Заключение
По крайней мере, три основных признаки отличают род Homo
- Вертикальная походка.
- Изготовление орудий труда.
- Развитый мозг и речь
- К пункту 1: В противоположность общепринятым мнениям из школьных учебников абсолютно изменилось наше представление о том, что австралопитеки имели, подобно человеку, вертикальную походку. Эксперты склоняются к тому, что австралопитеки представляли собой форму, обитающую больше на деревьях, чем на земле.
- К пункту 2: Ни одной из известных форм австралопитеков до настоящего времени невозможно с абсолютной уверенностью приписать использование орудий производства, хотя можно предположить, что, подобно современным шимпанзе, они могли располагать в своем комплексе поведения простейшими приемами использования палок и камней.
- К пункту 3: Величина мозга у австралопитеков не должна расцениваться как ключевой признак для определения его местоположения в цепи эволюции между животными и человеком. Получить достаточно веские доказательства наличия речевой способности у австралопитеков не представляется возможным.
Обилие гипотез о таксономическом и филогенетическом положении австралопитеков может несколько запутать читателя. михаель Дои, один из ведущих антропологов, резюмируя этот круг проблем, писал в 1985 году: "В общем таксономическое положение австралопитеков кажется таким же запутанным, как и ранее".
Некоторые признаки австралопитеков расцениваются как показатели процесса гоминизации (антропогенеза). Однако, весь комплекс признаков позволяет также сделать вывод о том, что перед нами специально приспособленная форма, обитавшая на деревьях. Авторы этого учебника присоединяются к мнению тех антропологов, которые не рассматривают эту высокоспециализированную вымершую группу приматов как предка человека.
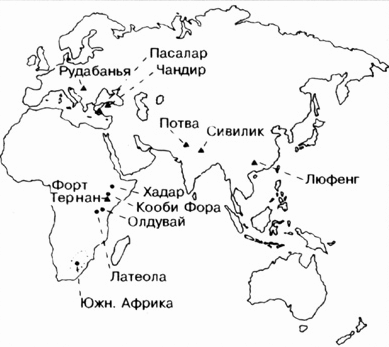
Рис. 9.6. Наиболее важные находки ископаемых приматов, относящиеся к эпохе миоцена (рамапитекообразные - места обнаружения обозначены треугольниками; австралопитеки - места обнаружения обозначены точками) и плиоцена.
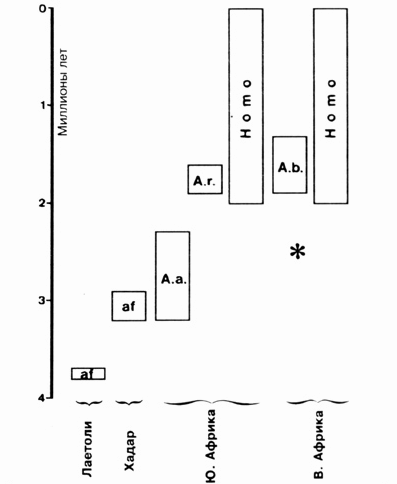
Рис. 9.7. Временное и географическое положение наиболее важных находок австралопитеков, которые иногда встречаются наряду с человеком. Географические места находок ископаемых экземпляров нанесены на рис. 9.6. Звездочка указывает на местоположение последней находки австралопитека, относящегося к гиперробустному австралопитеку (см. текст и рис. 9.8). А.а. - A. africanus; at - A. afarensis, A.r. - A. rohustus; A.b. - A. boisey
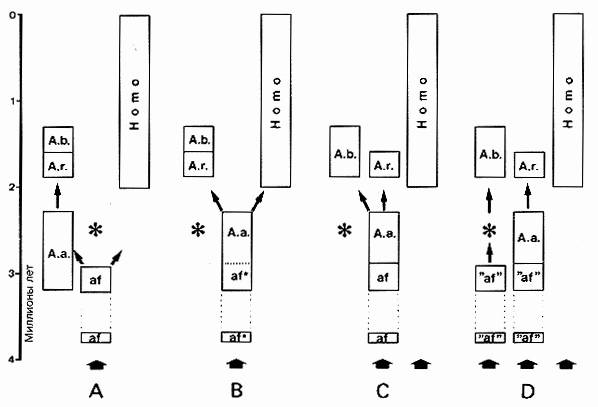
Рис.. 9.8. Три варианта родословного дерева видов Australopithecus и Homo (A - С) и родословное дерево, которое с 1986 года стало наиболее популярным, (вариант Д). Родословное дерево (вариант А), предложено johanson и white в 1979 г. ставит вид afarensis в непосредственное родство по восходящей линии, в то время как вид A. africanus вместе с формами австралопитеков robustus образуют побочную ветвь. Согласно варианту В в родословном дереве вид A africanus является непосредственным предком человека (вместе с формой robustus). В этом случае вид A. africanus (обозначен звездочкой) рассматривается либо идентичным виду africanus (tobias, 1980), либо его предком. Древняя линия развития человека, которая существовала параллельно с линией развития австралопитеков по крайней мере в течение 5 млн. лет и не ведет своего начала ни от одной из известных на сегодняшний день форм австралопитеков, изображена в варианте (" (leakey, 1981). Критика, направленная на местоположение вида A. afarensis справедлива для варианта В родословного дерева и, в еще большей степени, для варианта Д. Уже в 1981 г. палеонтолог Ольсон указал на наличие в ископаемом материале из Хадара двух форм австралопитеков: robustus и gracilis и выразил, наряду с другими учеными, сомнение в правомерности объединения их в один ряд ("af"). Эта схема родословного дерева позволяет легко подобрать место для недавно открытого вида австралопитеков hyperrobustus, обозначенного звездочкой, который датируется гораздо большим возрастом, чем все остальные (Walker, Ifaky 1986). Я то же время другие варианты родословных деревьев должны для этого в большей или меньшей степени подвергнуться изменениям.
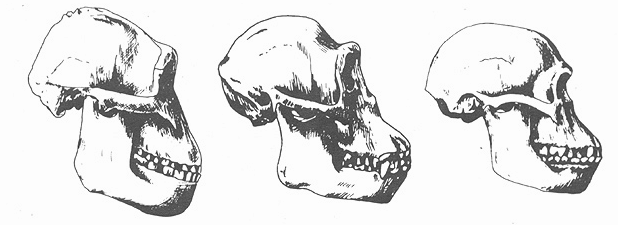
Рис. 9.9. Череп шимпанзе (посредине) к сравнении с черепами Australopithecus robustus (cлева) и A africanus (справа).
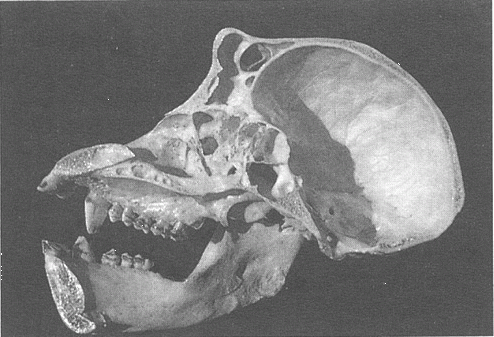
Рис. 9.10. Продольный разрез черепа шимпанзе. На внутренней стороне черепной коробки видны четкие отпечатки кровеносных сосудов и отпечатки мозговых извилин. Это помогает сделать некоторые выводы о морфологии мозга.
Таблица 9.1. Соотношение объема мозга с весом тела - наиболее распространенный показатель уровня развития и интеллекта. Однако при более близком рассмотрении это соотношение не является показательным. Иначе обезьяны-капуцины оказались бы на более высокой организационной ступени, чем человек. К таблице приводятся усредненные величины для индивидуумов мужского пола (по knussmann, 1980).
|
Вид |
Объем мозга см 3 |
Вес тела в г |
Отношение |
|
Человек |
1400 |
70000 |
1:50 |
|
Шимпанзе |
400 |
45000 |
1:110 |
|
Горилла |
530 |
175000 |
1:330 |
|
Орангутан |
430 |
75000 |
1:175 |
|
Гиббон |
100 |
6000 |
1:60 |
|
Капуцины |
75 |
2000 |
1:30 |



