




История происхождения и развития жизни
8.7.8. Эволюция лошадей
Из всех млекопитающих животных остановимся на лошадях, так как они являются наиболее распространенным примером. Почти в каждом учебнике, излагающем эволюционную теорию, можно прочитать, что история эволюционного развития непарнокопытных (Perissodactyla) из всех млекопитающих (и даже среди всех позвоночных животных) лучше и в большем объеме снабжена ископаемыми доказательствами. Их даже называют иногда "триумфальным конем" эволюции. К непарнокопытным относятся лошади, тапиры и носороги, а также другие формы, встречающиеся в ископаемом виде.
"Лошади утренней зари"
Все непарнокопытные должны были произойти от одного предка (Tetraclaenodon), форма, которая датируется палеоценом. Между этой формой и самыми древними непарнокопытными (Hyracotherium) не найдено, однако, никаких промежуточных звеньев. Ископаемые представители отряда Hyracotherium встречаются как в Европе, так и в Соединенных Штатах Америки, они назывались там "Eohippus" ("утренней зари"). Это животное считается первым общепризнанным предшественником современных лошадей (древняя "пралошадь") (рис. 8.74). Оно обладало на передних ногах четырьмя, а на задних ногах - тремя пальцами. Обнаруженный на руднике Мессель близ Дармштадта Propaleotherium (рис. 8.75) классифицируется как более близкий родственник Hyracotherium. (Общеупотребительное представление об эволюции лошадей коротко изображено на рис. 8.76, и мы рекомендуем сначала изучить этот рисунок).
Но действительно ли Hyracotherium являлся "пралошадью"? Будет поучительным узнать, что поначалу палеонтологи сравнивали эту находку с тапирами, носорогами, грызунами, даманами (Hyracoidea), а после сравнивали по внешнему виду с дукерами (хохлатыми антилопами), но не с лошадью. В учебниках распространено мнение о том, что от древнейших видов Hyracotherium произошли не только лошади, но и тапиры, и носороги, а также другие виды вымерших непарнокопытных. Таким образом, нельзя ответить однозначно на вопрос, почему Hyracotherium называют "пралошадью", а не "пратапиром" или "праносорогом". В действительности же известный специалист по эволюции млекопитающих животных из Вены эрих тениуг пишет, что у Hyracotherium отсутствуют типично лошадиные признаки. Известнейший современный исследователь эволюции лошадей Дж. Симпсон констатирует, что Hyracotherium больше похожи на даманов, также имевших спереди четыре, а сзади - три пальца на конечностях, чем на современных лошадей. Почему же тогда Hyracotherium попадает по классификации в семейство лошадей (Equidae) (romer, 1966, с. 386), а даман (Нутах) в совсем другой, свой собственный отряд? В этом вопросе мы хотели бы обратить внимание читателей, во-первых, что название "пралошадь" основывается не на анатомии ископаемого, а на предполагаемых филогенетических взаимосвязях.
Речь идет в данном случае о названии, с помощью которого внушается эволюционная взаимосвязь. Этому же служит явно выраженное стремление некоторых палеонтологов более раннее (и, таким образом, по правилам номенклатуры законное) наименование поменять на более позднее "Eohippus" (что может быть переведено приблизительно, как "лошади утренней зари") с тем, чтобы "названием Hyracotherium не вводить в заблуждение неподготовленного читателя" (davies, 1977, с. 54, из nilsson, 1954, с. 548). Cимпсон выразил свое мнение, подчеркивая, как "неудачно" выбрано название Hyracotherium. Мы касаемся так подробно этой тенденции, чтобы продемонстрировать, как предвзятое мнение, иногда вопреки очевидным фактам, может повлиять на биолого-таксономическую направленность мышления.
Зигзагообразная эволюция?
В начале исследований процесс эволюции лошадей, казалось, можно было толковать однозначно и прямолинейно, однако, с накоплением данных результатов раскопок ископаемых положение становилось все более сложным, и из родословного дерева образовался целый родословный "кустарник" с многочисленными побочными ветвями и линиями, обозначающими вымершие группы. Следует предположить, что развитие предполагаемых предков лошадей между Америкой и Европой шло извилистым путем и что были, вероятно, периоды вымирания. Кроме всего прочего, следует признать, что отдельные признаки (величина, зубы, пальцы на ногах), во-первых, не всегда развивались в "желаемом" направлении и, во-вторых, развивались в различных формах независимо друг от друга. Так, например, Neohipparion, трехпалая форма с челюстью травоядного, появляется рядом с Pliohippus (однопалое, имеющее также челюсть травоядного, см. рис. 8.76). Как же это возможно, если однокопытная форма имеет для обитания в степи явное селективное преимущество перед трехпалой формой? Имеются и такие признаки, которые совсем не вписываются в предложенную эволюционную схему, - здесь следует назвать такой признак, как количество ребер, которое регулярно колеблется от 15 до 19. (Можно возразить, что даже при планомерном выведении пород число ребер колеблется в гораздо меньшем масштабе, и поэтому не может служить достаточным доказательством. Однако именно этот факт приводится все время в качестве серьезного доказательства возможных несоответствий). Подобные факты дали повод авторам говорить о "зигзагообразной" эволюции, иногда даже эволюционный ряд лошади привлекают в качестве доказательства обратимости эволюции.
Отсутствующие переходные этапы
Итак, переходы между "станциями" на предполагаемом эволюционном пути лошади представляют собой отнюдь не непрерывный сплошной путь: на нем можно обозначить скачкообразные этапы. Прогрессирующее увеличение можно наблюдать не всегда, так как Orohippus, Archaeohippus и Nannippus были меньше по размерам, чем их предполагаемые предки. Из всех форм как раз самая древняя и является самой большой по величине (величина до подмышечной впадины 30 см). Следующий род Mesohippus скачкообразно увеличил свой рост приблизительно на 60 см (до подмышечной впадины), промежуточные формы неизвестны. Плавного перехода от Miohippus к Merychippus (первая форма питалась листвой, вторая - травоядная форма) тоже не наблюдается, хотя иногда к переходной форме причисляют Parahippus (рис. 8.76). Однако это животное обладало формой челюсти, характерной для форм, питающихся листвой, и строением стопы, приспособленной для лесных условий. Поэтому переход к челюсти Merychippus, характерной для травоядных, остается без промежуточного звена. И эволюция высоты коронок также не представляется нам плавно протекавшим процессом, и здесь (например, от Merychippus до Pliohippus) наблюдаются скачки именно там, где, согласно эволюционной теории, мы должны наблюдать многочисленные плавные переходы (рис. 8.78). Во всех схемах родословного дерева, приведенных в учебниках, бросается в глаза, что в процессе "редукции" (обратного развития) количества пальцев на ногах имеются скачки (рис. 8.79); в процессе развития от Hyracotherium к Mesohippus без перехода и какого-либо промежуточного звена исчезает четвертый палец на передней ноге - основания для этого, неизвестны. От Merychippus к однокопытной (непарнокопытной) истинной лошади Pliohippus происходит редукция стопы от четырехпалой до однопалой (однокопытной) стопы "с достаточно большой скоростью" (simpson, 1977, с. 199), другими словами, с перерывами и без истинно промежуточных форм. Относительно эволюции ноги и стопы Симпсон пишет в заключении (1970, с. 203), что это развитие нельзя рассматривать как "широкую проторенную дорогу от Eohippus к Equus", но, напротив, как серию четко разграниченных, явно различающихся стадий".
У вымершего южноамериканского отряда непарнокопытных Litopterna обнаруживается странным образом обратное предполагаемому по родословному древу развитие: Thoatherium (нижний миоцен) имеет однокопытные конечности, род такого же возраста Diadiaphoras - ослабленные боковые пальцы на стопе (почти как у Merychippus), a y Macrauclienia - формы ледникового периода - наблюдается полностью сформированная трехпалая стопа (сравнимая со стопой Mesohippus). Неужели у Litopterna, несмотря на увеличение в росте, произошла эволюция от "высокоспециализированной" однокопытной стопы к "более примитивной" трехпалой? Если даже переместить Thoatherium и Diadiaphorus в другое семейство, чем Macrauclienia, и не предполагать прямолинейного происхождения Litopterna непарнокопытных животных друг от друга, то остается ЯВНЫМ, ЧТО более ранние (третичный период) формы представляют собой высокоспециализированных однокопытных, в то время как более поздние (ледниковый период) виды имеют полностью функционирующие трехпалые, и таким образом, неспециализированные конечности. Конечно, можно в рамках эволюционной теории обосновать и эту тенденцию в развитии непарнокопытных южноамериканских, противоречащую тенденциям развития лошади. Ведь в эволюционной теории речь всегда идет о гипотетических строениях, с помощью которых могут быть впоследствии объяснены все мыслимые направления развития. Так может ли это вообще служить аргументированным доказательством?
В таблице 8.4 перечислены 7 групп признаков, которые при изучении эволюции лошадей от животного, обитающего в лесах и питающегося листвой, до травоядного обитателя степей необходимо учесть и проконтролировать в динамике их изменения. По первым пяти пунктам палеонтологическое наследие не может дать последовательных убедительных доказательств, и этот факт заслуживает тем большего к себе внимания, чем громче сторонники эволюционного учения возводят "на пьедестал" в качестве "триумфа эволюционного учения" теорию эволюции лошади.
Рассматривая проблему в общем и целом, риппель пишет (1983, с. 74), что "только начальные стадии Hyracotherium и Mesohippus в Северной Америке могут представлять собой до некоторой степени последовательный процесс развития. Более поздний этап развития лошади должен быть поделен на множество независимых линий эволюционного развития, с периодически проявляющимися тенденциями обратного хода развития, которые могли бы оправдать противоречивость развития признаков" Симпсон выражает мнение, что, "вероятно, не существует ни одного признака, который бы развивался постепенно в русле единой тенденции у всех видов лошадеобразных в равной степени одним и тем же образом" (1944, с. 159).
Несмотря на контраргументы, приведенные выше против эволюционного ряда, ведущего к современной лошади, можно, исходя из постулатов эволюционной теории, классифицировать и выстроить в ряд все обнаруженные учеными ископаемые. Мы же, однако, старались довести до вашего сведения, что этот ряд имеет достаточно весомые провалы, и банк данных ископаемых никоим образом не является банком бесспорных аргументов в пользу этого ряда.

Рис. 8.74. Так называемая "пралошадь" Hyracotherium. Эти животные считаются признанными предками лошадей. (Земельный музей природы, Карлсруэ.)
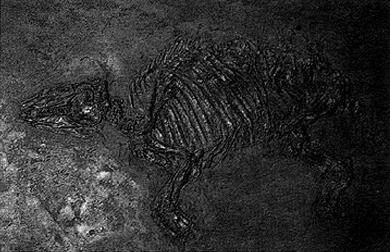
Рис. 8.75. "Пралошадь" Propaleotherium, относящаяся к эпохе эоцена, найдена на руднике Мессель близ Дармштадта. Предполагается, что это не прямой предок лошадей, но близкий родственник "пралошади". Подмышечная впадина около 30 см. (Земельный музей природы, Карлсруэ.)
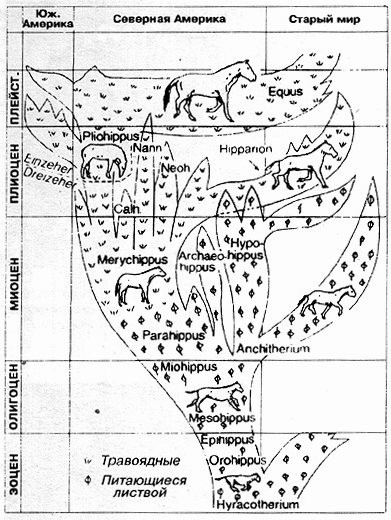
Рис. 8.76. Классическое родословное дерево лошадей. У Hyracotherium предполагается постоянное увеличение размеров, от кошки до лисы, до размеров современной лошади. Параллельно с этим должно было происходить сокращение числа пальцев на ногах при прохождении промежуточных ступеней, при этом грифельные косточки лошадей рассматриваются как утратившие свое функциональное значение. "Пралошади", судя по их челюстям (низкие коронки зубов) питались листвой, а при перестройке зубов на высоко коронный тип, по общему мнению, стали травоядными. Пытаются связать соответствующие изменения с давлением отбора в направлении дальнейшего развития как на мягкой лесной почве, так и на жесткой степной (современные лошади, как однокопытные), а также провести связь с питанием, соответствующим ареалу обитания. Недавно палеонтологам Естественно-исторического музея Зенкенберга во Франкфурте удалось доказать на основании изучения содержимого ископаемого желудка "пралошади", обнаруженной на руднике Мессель (Propaleotherium), что это животное - как и следовало предполагать, судя по состоянию зубов, -действительно питалось листвой. (По Thenius, 1981, по Simpson.)

Рис. 8.77 Некоторые представители лошадей . Справа на лево Hyracotherium, Mesohippus, Merychippus, Pliohippus, зебра Грэви. ( Земельный музей природы Карлструэ)
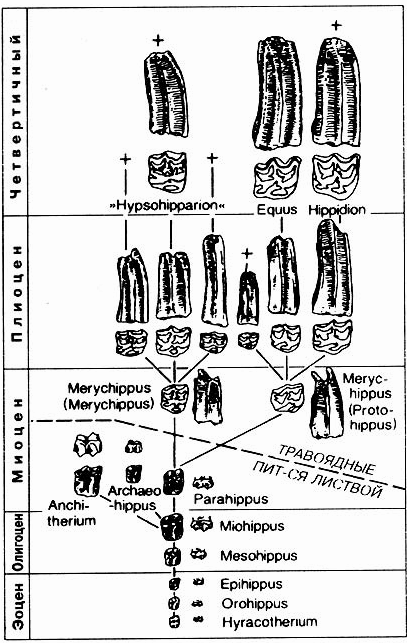
Рис. 8.78. Эволюция зубов лошади согласно классическим представлениям. Четко обозначенная граница между зубами с нижними коронками, характерными для животных, питающихся листвой (до Parahippus), и зубами с высокими коронками, характерными для травоядных (от Merychippnus). (По Thenius, 1969.)
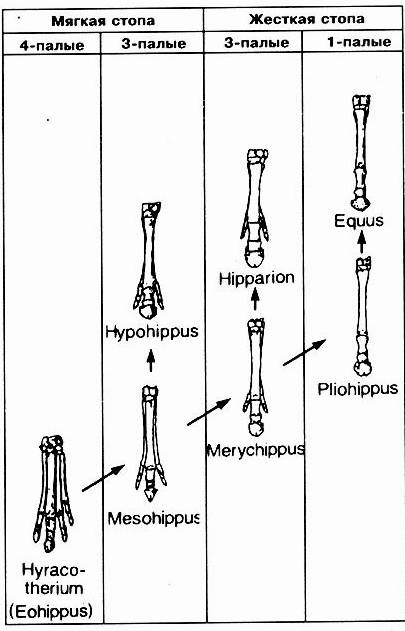
Рис. 8.79. Эволюция стопы (передней конечности)
лошади в рамках классических представлении. Каждая колонка представляет определенный
механический тип. Плавные переходные стадии между отдельными типами отсутствуют.
(По .Simpson, 1977)
|
Таблица 8.4. Требуемые изменения нескольких групп признаков в постулируемой эволюции лошади. Признаки, перечисленные в пунктах 6 и 7, в принципе, не могут быть доказаны с помощью ископаемых. |
|
1. Возникновение нового рисунка складок зубной эмали |
|
2. Смена зубов с низкими коронками, характерных для животных, питающихся листвой деревьев, на чубы с высокими коронками, характерные для травоядных. |
|
3. Создание механизма, который на протяжении всей жизни лошади помогает сохранять высоту чубов на одном и том же уровне (таким образом, как бы "выталкивает" их из челюсти на требуемую высоту), деформирующемся мри питании растительной пищей (травой). |
|
4. Возникновение абсолютно новой зубной субстанции - зубного цемента. |
|
5. Постепенная редукция пальцев на сгоне с параллельно проходящими изменениями строения стоны: от стопы, приспособленной к мягкой лесной почве, до стопы, приспособленной к жесткой почве степи. |
|
6. Возникновение ноною аппарата и пищеварения. |
|
7. Возникновение нового характера поведения. |



