




История происхождения и развития жизни
6.5.2. Основные правила биогенетики
В 1866 году Эрнст Геккель сформулировал в качестве основного биогенетического закона положение о том, что человек на стадиях раннего эмбрионального развития (в онтогенезе) повторяет в сжатом виде всю историю развития рода (филогенез): "Развитие всякого организма есть ускоренное повторение его филогенеза". Утверждали, например (а иногда утверждают еще сегодня), что в эмбриональном развитии человека присутствуют стадии развития рыб и амфибий. Значит, в нем можно обнаружить признаки животных и тем самым становится ясен филогенез человека в ходе его онтогенеза. Этот закон был распространен также и на другие организмы, и в общей форме в нем оговаривается, что организмы во время своего эмбрионального развития проходят все стадии исторического развития. Полагали также, что в эмбриональных стадиях живущих ныне животных и растений можно наблюдать организационные признаки филогенетического развития древних существ: образовавшиеся в эмбриональном состоянии и претерпевшие затем обратное развитие органы представляют собой бесполезные остатки прошлого, которые можно объяснить только происхождением живых существ друг от друга. Сам Геккель рассматривал свой основной биогенетический закон как доказательство учения о происхождении. Он писал (1920, с. 236): "В тесной взаимосвязи эмбри- и филогенеза усматриваю я одно из важнейших и неопровержимых доказательств эволюционной теории".
К истории разработки биогенетического правила
Когда Геккель сформулировал основной биогенетический закон (он говорил исключительно о законе), у него в распоряжении имелось множество данных для его обоснования. Человеческие эмбрионы на ранней стадии развития ничтожно малы и абсолютно прозрачны, так что их нормальным способом наблюдать невозможно. Они настолько подвержены тлению, что легко гибнут, и без особых методов фиксации их нельзя сохранить от угрозы быстрого наступающего сразу же после гибели распада. Но во времена геккеля не было подходящих приборов а-ля фиксации, равно как и хороших гистологических методик окрашивания, которые бы позволили сделать видимыми нежнейшие структуры молодых эмбрионов и дать, таким образом, более точные сведения об эмбриональном развитии человека. Лишь в 1950 году были изучены молодые человеческие яйцеклетки. Плохо фиксированные препараты послужили причиной того, что Геккель не имел возможности подтвердить свое мнение убедительными доказательствами. Так как в распоряжении ученого находились уже испорченные по форме и строению зародыши (следовательно артефакты), то Геккель мог констатировать лишь поверхностное сходство, из которого он впоследствии сделал ложный вывод об истинном сходстве.
Кроме того, грекклю было предъявлено обвинение, что он фальсифицировал снимки эмбрионов. Он признался ("Berliner Volkszeitung" от 29.12.1908): "... Хочу начать сразу же с покаянного признания, что часть, малая часть моих многочисленных снимков эмбрионов действительно подделана, та в которой предложенный для обследования материал был слишком неполный, недостаточный; для восстановления связной цепи развития необходимо было заполнить провалы в ней гипотезами". Совершенно ясно, что Геккель руководствовался в своих исследованиях скорее идеей, чем фактами. Но сегодня для нас не имеет никакого значения, какими подходами в своих исследованиях пользовался Геккель, хотя его идеи и сегодня все еще дискутируются.
Утверждения Геккеля были тем временем опровергнуты. Представления о том, что человек был сначала "наполовину животным" и лишь в ходе эмбрионального развития превратился в настоящего человека, не выдерживают натиска многочисленных данных исследований эмбрионов человека. В научной литературе на эту тему сформулированный геккелнм основной закон биогенетики более уже не обсуждается. Но, несмотря на это, большинство биологов продолжает считать это основным биогенетическим правилом, имеющим множество исключений. Во всяком случае, в школьных учебниках и в популярной литературе этот закон изучается в его прежнем устаревшем виде. Поэтому, мы сначала приведем доказательства того, что формулировка Геккеля неприемлема.
Палингенез и ценогенез
Предположение о том, что в онтогенезе происходит полное повторение филогенетического развития, опровергнуто многочисленными примерами. Многие насекомые проходят в своем развитии стадию куколки, во время которой они не получают питания, неподвижны и, тем самым, относительно незащищены. Эта стадия не может соответствовать ни одной предыдущей стадии развития взрослого насекомого. Исходя из этого, никто не возьмется утверждать, что его предки имели во взрослом состоянии желточный мешок, хотя этот мешок, как и все остальные эмбриональные образования, можно наблюдать в зародыше. Важным признаком раннего эмбрионального развития у человека являются сильно развитые области головного и спинного мозга, которые у более взрослого эмбриона составляют большую часть их объема. Такое рапное развитие головного и спинного мозга рассматривается как рекапитуляция, свидетельствующая о том, что наши предки, вероятно, обладали большей мозговой субстанцией, чем мы. Подобные примеры встречаются на каждом шагу.
Поэтому, следует различать палингенетические и ценогенетические признаки. Палингенетическими (гр. palin = назад) называются признаки, которые должны повторять стадии развития предполагаемого предка, ценогенетическими (гр. kainos = новый) - такие, по которым нельзя определить связь с предполагаемым предком в филогенезе. Перечисленные выше признаки являются примерами ценогенеза.
По онтогенетическим данным нельзя судить о филогенезе, "поскольку, только зная филогенез, можно решить, что именно происходит на определенной стадии онтогенеза - ценогенез или палингенез" (siewing. 1982, с. 271). Это. поясняется примером индивидуального развития гориллы, приведенном на рис. 6.19. Если применить в этом случае основное биогенетическое правило, то мы придем к выводу, что у горилл были человекоподобные предки, так как череп молодой гориллы имеет сходство с черепом человека.
С другой стороны, человек и макака-резус к началу их онтогенетического развития демонстрируют явно различные исходные структуры (рис. 6.20), которые лишь позднее приобретают сходство.
Даже самые первые онтогенетические фазы не могут соответствовать предполагаемому филогенезу, так как в процессе филогенеза первые одно- и малоклеточные организмы должны были иметь другой способ питания (через пищеварительный тракт), чем развивающиеся зародыши во время онтогенеза (деление имеющегося в яйцеклетке материала, питание через плаценту).
Раннее эмбриональное развитие вплоть до стадии гаструлы протекает у разных классов позвоночных животных (рыб, амфибий, рептилий, птиц и млекопитающих) во многом различно и абсолютно вопреки основному биогенетическому правилу. И лишь следующие фазы указывают на конвергентное развитие в известных схожих друг с другом стадиях ("Закон эмбрионального сходства позвоночных животных"). Это означает: основы строения тела позвоночных животных закладываются абсолютно разными способами. Последнее утверждение приближает нас к выводу, что причины имеющихся путей развития следует искать не в предполагаемом процессе филогенеза, а в функциональных потребностях.
По этим и другим причинам "основное правило биогенетики" сегодня ограничивается следующим утверждением: лишь части стадий онтогенеза являются рекапитулятивными. С точки зрения эволюционной теории лишь некоторые структуры рассматриваются как "память'' о предыдущих стадиях историко-родового развития. Поэтому в таком случае употребляют термин "рекапитуляционное развитие" отдельных органов или признаков. Об утрате функции пока речь не идет.
Если и пробовать установить филогенетическую связь между несколькими складками в области шеи и головы человеческого эмбриона с жаберными дугами рыб. то только потому, что предположение наличия подобных связей очень хорошо вписывается в контекст эволюционно - теоретических объяснений, который сложился и без того (см. Peters, 1975), но не потому, что они не имеют никакой функции или что это по - другому объяснить невозможно.
Бессмысленные ходы в развитии, которые указывают нам только на филогенез, должны быть исключены под воздействием отбора, так как они предполагают бесполезный расход времени и материала и, следовательно, являются селективно-отрицательными (см. 3.3).
Очень поучительным примером функциональности временно образованных структур является хорда (опорная трубка в области позвоночника) позвоночных животных. В ходе эмбрионального развития позвоночных животных до появления позвоночного столба на его месте сначала образуется хорда, которая затем деградирует, с точки зрения эволюционной теории это расценивается как доказательство того, что позвоночные животные произошли от определенных хордовых, которые во взрослом состоянии имеют хорду в качестве опорной трубки (а не в качестве позвоночного столба). Гутман и Боник (1981, с. 108) писали, однако, о функции хорды у позвоночных следующее: "Несмотря на то, что хорда у многих позвоночных животных в период эмбрионального развития выражена лишь тоненькой линией клеток, она является необходимым, неотъемлемым органом. Так, например, среди мышей известны мутанты, у которых развивается слишком тонкая хорда. Последствием этого является срастание многих позвоночных сегментов, так как остались неразвитыми межпозвоночные хрящи... Более убедительного доказательства пользы от столь ничтожно малого "исторического остатка" не существует". Другой пример: зачатки зубов для беззубых млекопитающих необходимы для правильного формирования челюстей. Таким образом, польза от сформированных в онтогенезе лишь на некоторое время органов очевидна. И далее становится понятно, что органы, чтобы правильно оценить их значение, никогда нельзя рассматривать изолированно.
Впрочем, уже сам Геккель, по существу, противоречил своему собственному закону: "Точное повторение филогенетического в бионтическом развитии извращается и вытесняется вторичной приспособляемостью" (по peters, 1975).
Заключение: Решить, что обусловлено филогенетическим развитием (палингенез), а что с ним не связано (ценегенез), представляется возможным лишь при признании преимущества эволюционной теории и предполагаемого процесса эволюции. Это означает, однако, что данные онтогенеза не могут считаться независимыми доказательствами эволюции.
Данные исследований эмбриологии человека
Представление об органах, утративших свои функции в человеческом организме, было опровергнуто проведенными тщательными исследованиями. Только если рассматривать органы эмбриона вне рамок индивидуального развития, то можно изображать их некоторые образования в качестве рудиментов или атавизмов. "Во время онтогенеза во многих случаях сначала образуются признаки, присущие предкам, которые не имеют значения для жизни вида" (DIEHL, 1980, с. 55). Только реконструировав историю происхождения, можно легко объяснить и полностью понять рудиментарные органы. На самом деле не существует ни органов, потерявших свои функции, ни атавизмов. Каждый орган в рамках формирования в процессе роста является конструктивно необходимым. Это можно подкрепить несколькими примерами, которые общеизвестны как "филогенетическая память". Конструктивную необходимость можно доказать на эмбриональном уровне, если тщательно изучить последовательность разных стадий развития, формирование так называемых зародышевых листков, путем дифференциации которых происходит закладка органов и тканей. При этом вопрос о цели процессов развития (зачем?) остается вне рассмотрения. Его значение для послеродового развития не должно нас поначалу интересовать. Для тех, кого интересуют подробности, мы рекомендуем дополнительную литературу.
Необходимое знание деталей можно получить, изучая собрание документальных данных исследований человеческих эмбрионов, произведенных биологом Блехшмидтом (Анатомический институт университета г. Геттинген, рис, 6.21), Оно состоит из 64 муляжей высотой один метр, представляющих эмбрионы человека в течение первого и второго месяца развития. Эта коллекция составлялась и тщательно выверялась в течение 30 лет. В основе ее лежат 200000 отдельных гистологических препаратов эмбрионов. Это собрание экспонатов является всемирно признанным и вошло в международные каталоги (blechschmidt, 1973).
Образование органов, которое кажется оправданным только филогенетически
"Жабры"
Часто можно встретить утверждения, что у человеческих эмбрионов в первые месяцы жизни наблюдаются четыре жаберных дуги, которые впоследствии исчезают. "Выходящие из сердца сосуды проходят точно так же, как ведущие к жаберным дугам кровеносные сосуды у рыбы; позднее они претерпевают обратное развитие. Иногда открытой остается даже одна складка, соответствующая жаберной складке у взрослых в виде так называемой горловой фистулы" (knodel и bayrhuber, 1983, с. 385).
Жабры - это органы дыхания, характерные особенно для рыб. У человека не планируется никаких жабер, не образуются они и как промежуточное звено. Человеческий эмбрион величиной в 3,4 мм в передней части своего тела изгибается. Изгиб возникает как извилина роста. Он выражается в характерных складках изгиба между лбом и сердечным утолщением в области, в которой у эмбриона на ранней стадии развития находится лицо. Извилины роста имеют следующую динамику: эмбриональная нервная трубка растет особенно сильно в длину - это означает, что она является основным потребителем питания. Во-первых, идущий от сердца (парный) кровеносный сосуд подает 'для этого необходимое питание, хотя сам не растет в такой же мере, как нервная трубка. Тем самым, зачаток аорты по всей своей длине оказывает сопротивление процессу роста связанной с ним тканями нервной трубки, и она изгибается на своем свободном конце (в области головы) над утолщением в области сердца. При этом возникают складки изгибов как в области внешней, так и внутренней стенки корпуса (рис. 6.22 - 6.25). Складки изгибов возникают как внутренние дуги, которые обхватывают поперек головную кишку: 1. дуга нижней челюсти; 2. подъязычная дуга, затем две дуги гортани. В них впоследствии развивается скелет нижней челюсти, подъязычной кости и гортани.
С ростом эмбриона внутренние дуги становятся все больше, а ткани в них натягиваются. Это служит структурной основой для нервов и кровеносных сосудов.
Эти кровеносные сосуды представляют собой короткие перемычки (анастомозы) между сосудом, по которому кровь вытекает из сердца, и парным зачатком аорты. От изгиба эмбриона зависит интенсивность кровоснабжения: сердце пытается с каждым ударом пульса вытеснить кровь по самому короткому пути. Сначала, когда изгиб начинает формироваться, это происходит через первый сосуд внутренней дуги. С увеличением изгиба дуги значение первого кровеносного сосуда снижается. Затем особое значение приобретает второй кровеносный сосуд внутренней дуги для оттока крови (напрямую), вскоре эта функция переходит к третьему, затем к четвертому сосуду. Первые три сосуда с их извилистым оттоком от основного сосуда постепенно преобразуются, частично претерпевая обратное развитие. Короткая перемычка между аортами остается открытой до тех пор, пока легочное кровообращение после рождения не исключит ее полностью и вся кровь не будет протекать через сосуды легких.
С формой и функцией жабер складки сгиба у человека не имеют ничего общего. Они в большей степени являются следствием раннего периода возрастного изгиба головы, связанного с интенсивным ростом головного мозга относительно кровеносных сосудов. Между складками изгиба образуются складки, в глубине которых внешние стенки корпуса вплотную прижаты к внутренним.
В области соприкосновения эктодерма и энтодерма могут быть настолько тесно прижаты друг к другу, что ни один подающий питание сосуд не может там проходить и клетки местами гибнут. Таким образом, возникают дефекты, которые болезненным образом в качестве складочных образований могут сохраняться вплоть до момента рождения (так называемые горловые фистулы). Они представляют собой аномальные разрывы на обычно гладкой стенке тел, а вовсе не открытые жаберные складки. Этот же принцип развития подобных отверстий (коррозии) срабатывает и в других областях тела в ходе их развития (ротовая и анальная мембрана, присоединение почечных канальцев к почечной лоханке и т. д.).
Эволюция млекопитающих должна была проходить от рыб через стадию амфибий и рептилий. Но ближайшие к рыбам амфибии не обладают ни в одной из стадий развития жаберными щелями, ни даже жаберными бороздками (лягушки и тритоны имеют сначала внешние, затем внутренние жабры и, наконец, легкие). Поэтому нам представляется абсурдным считать бороздки в области шеи и лица человеческого эмбриона "жаберными щелями".
"Хвост"
Отмечающееся иногда наличие в редких случаях у новорожденных "хвостиков" расценивается как дальнейшее доказательство филогенетического развития.
Кроме того, доказано, что в эмбрионе имеются зачатки хвоста вместе с хвостовой частью позвоночника.
К сожалению, часто нижний заостренный конец тела ошибочно принимается за, так называемый, хвост. У человека, однако, ни в одной из стадий не образуется ни хвоста, ни даже позвоночного столба хвоста. В противоположность верхней части, мало сдерживаемой в росте, нижняя часть остается, вследствие относительно низкого уровня кровоснабжения, тонкой и заостренной (рис. 6.26 и 6.27).
У эмбриона размером в 10 мм конусообразный спинной мозг вместе с растущим головным мозгом втягивается в канал соединительной ткани и при этом стягивается в кожной трубке назад к голове так, что нижний конец тела ослабевает. Направление расположения нервов у взрослых иллюстрирует этот основной процесс созревания плода на протяжении всей жизни (диафрагма, например, обеспечивает свою жизнедеятельность с помощью шейных нервов). Таким образом, нижний конец тела человеческого эмбриона уменьшается, а не удлиняется в виде хвоста. Сравнительные измерения показали, что на нижнем конце тела человеческого эмбриона не происходит удлинения, похожего на хвостовое образование.
Вследствие отсутствия спинного мозга в области нижнего конца тела отсутствует здесь и хорошо сформированный позвоночный столб. Известно, что позвоночный столб возникает в тесной конструктивной взаимосвязи с развитием спинного мозга. Крошечный копчик есть следствие слабого развития спинного мозга в нижней части тела эмбриона.
Чрезвычайно редко встречающиеся хвостообразные выросты, возникшие в процессе эмбрионального развития, не являются наследственными. И не было случая, чтобы подобное кожное образование из соединительной ткани содержало хотя бы фрагмент позвоночного столба, который обязательно присутствует у всех обладающих хвостами позвоночных животных. К тому же эти "хвостики" располагаются в большинстве случаев "не на положенном" месте встречаются дополнительно также и у хвостатых млекопитающих.
"Плавательные перепонки"
Мнение о том, что у человека в качестве промежуточного звена развиваются плавательный перепонки, является следствием неправильного истолкования этапов эмбрионального развития человеческой руки.
Возможное возникновение так называемой "плавательной перепонки" являлось, вероятно, пограничным случаем нормы, а не филогенетической рекапитуляцией. Зачатки пальцев удлиняются благодаря преимущественному росту со стороны своих окончаний (рис. 6.28). При этом кровеносные сосуды и нервы в области тонких лучевидных образований на ладони развиваются в соответствии с развивающимся скелетом пальцев. Если это соответствие в развитии в каком-то случае, отклоняясь от нормы, будет несколько меньшим, то между пальцами может возникнуть маленькая кожная - складочка.
Ушные мышцы
В результате некоторой тренировки многие люди могут с. помощью специальных мускулов заставить уши двигаться самостоятельно. Все мускулы возникают в так называемых зонах растяжения. Таков принцип развития. И так называемые "рудиментарные" ушные мышцы не являются в этом случае исключением Они возникают в результате роста ушной раковины в области растяжения, когда (у пятнадцатимиллиметрового эмбриона) в ходе дальнейшего развития затылок относительно ушной раковины развивается назад и вверх. В процессе роста эмбриона они конструктивно необходимы, но после этого периода теряют свое значение.
Первичный и вторичный челюстной сустав
Образование первичного и вторичного челюстного сустава (сустава слуховых косточек и сустава верхней и нижней челюстей), вероятно, можно объяснить с помощью гомологии. "Первичный" и "вторичный" означают не перестройку одного и того же сустава, но образование различных суставов во временной последовательности. Направление развития в онтогенезе состоит в этом случае в следующем: сначала основание черепа и меккелев хрящ (скелет нижней челюсти) представляли собой органичное единство. Но как только начинается относительное уменьшение площади, занимаемой сердцем, освобождается место для развития области лица, удлиняется скелет нижней и верхней челюсти, и хрящ как бы "опрокидывается" вниз. При этом возникают "разломы" (переломы роста) между хрящом и основанием черепа (первичный челюстной сустав со слуховыми косточками). Этот процесс оказывает влияние на направление развития хряща, угол отклонения которого от основания черепа все увеличивается (рис. 6.29). Только лишь минерализованный хрящ в основании черепа представляет собой истинный (так называемый вторичный) челюстной сустав (сустав между костью нижней челюсти и височной костью). Если бы в данном случае в онтогенезе имело бы место лишь повторение филогенеза, то предполагаемые предки животных должны были сначала лишь обладать способностью слышать и только затем еще и способностью питаться.
Сегментация стенок тела (метамерия)
Так как возникновение метамерии человеческого тела не было разъяснено, ученые полагали, что можно найти выход в сравнении с животным миром (например, с рыбами) и тем самым получить достаточно полное толкование этого явления. С точки зрения динамики роста, метамерия является следствием процесса жизнеобеспечения сосудов спинного мозга, происходящего в определенные, регулярные промежутки времени, то есть следствием процесса его интенсивного роста (рис.6.30, слева). Даже в человеческом эмбрионе, едва достигшем 2,5 миллиметровой величины, тело в области выхода сосудов, идущих от аорты к спинному мозгу и остающихся относительно небольшими по сравнению с величиной области, обеспечивающей их жизнедеятельность, сдерживается в росте своей поверхности. Поскольку сосуды соединены тканями с поверхностью тела, то препятствование росту сосудов вызывает некоторое втяжение стенок тела в области спины так, что еще не дифференцированная соединительная ткань в области спины подразделяется на сегменты (рис 6.30, справа). С удлинением спинного мозга и связанным с этим раздвижением названных выше сосудов первичные сегменты тела постепенно увеличиваются: в них появляются продольные плоскость растяжения, в которых развивается сегментная мускулатура спины. Поэтому сегментация является необходимым следствием происходящих процессов развития (конструктивное значение для роста сосудов спинного мозга).
"Пушок"
Если младенец появляется на свет с более интенсивным волосяным покровом, чем обычно, то это является пограничным с нормой случаем, так как все дети имеют в утробный период достаточно развитый волосяной покров.
Кожа человека в эмбриональный период имеет два слоя: один, повернутый в сторону околоплодных вод, растущий относительно медленно, и второй, прилегающий к питательному раствору, развивающийся значительно интенсивнее. Следствием неравномерного развития обоих слоев кожи является то, что, с основной стороны клетки размножаются быстрее (чем возле околоплодных вод) и, увеличиваясь в глубину, образуют нечто наподобие лесенки: зачатки волос. С возрастанием сопротивления росту со стороны окружающих их соедини -тельных тканей волосяные луковицы выталкивают волос наружу. В ходе эмбрионального развития волосы подвергаются обычному обмену веществ и впоследствии отторгаются, либо, соответственно, заменяются. Предполагают, что с одной стороны, белок, входящий в состав волос, служит для питания, а с другой - волосы сами по себе выполняют роль наполнителя кишечника и тем самым служат возбуждению перистальтики
Lanugo (лат.) - Первичный волосяной покров человеческого зародыша (он обычно исчезает за несколько недель до нормальных сроков рождения) не может являться рекапитуляцией волос предположительных предков человека - обезьян. Это заключение следует из того факта, что обезьяны (гориллы, шимпанзе, орангутаны) точно так же имеют Lanugo.
Грудные соски
В качестве следующего рудиментарного органа рассматривают грудные соски у индивидуумов мужского пола. Уже у человеческогo эмбриона 3,4 миллиметра величиной явно прослеживается наличие так называемого эктодермального кольца - некой уплотненной кольцеобразной зоны кожи, которая наиболее отчетливо видна ниже лица, на боковой стороне тела (что приблизительно соответствует молочным бугоркам многих животных). Это свидетельствует об уменьшенном росте участка кожи относительно бистро разлипающейся поверхности тела в области спины и живота. С достижением достаточной степени изгиба эмбриона в эктодермальном кольце появляются зоны с особенно ярко выраженной задержкой роста этих участков, на которых .эпителий образует так называемые "соски". Появляющиеся время от времени многочисленные соски могут возникать на изгибе этого кольца, так как именно там в наибольшей степени тормозится рост эктодермы. Подобные "соски" можно получить экспериментально путем местного ограничения роста эпителиальных участков.
"Желточный пузырь"
Термин "желточный пузырь" вызывает зачастую неправильные ассоциации. Но именно он является конструктивно необходимым в процессе роста у человека. Его зачатки можно обнаружить уже в трехдневном зародыше в виде крошечной капельки жидкости. В двухкамерном яйце (приблизительно па пятый день) образуется верх желточного пузыря - внутренний зародышевый листок (энтодерма) человеческого эмбриона и дно амиона - внешний зародышевый листок (эктодерма). Желточный пузырь, благодаря своей тесной связи с материнским источником питания, выполняет в качестве аккумулятора диффузионных потоков функцию своего рода питательной железы. При этом питание клеток стенки желточного пузыря настолько интенсивно, что клетки здесь округляются, покидают это место и затем внедряются в молодой организм в качестве кровяных клеток. Таким образом, пока нет костей и костного мозга, желточный пузырь служит местом формирования кровяных клеток.
Заключение. Все так называемые рудиментарные органы или органы, появляющиеся лишь на каком-то определенном этапе ("рекапитуляционное развитие") имеют без исключения в рамках эмбрионального развития функционально возрастное назначение и при дифференциации последовательности фаз являются конструктивно необходимыми.
Рефлексы
На примере рефлексов должно стать еще более понятным, что естествоиспытатель может объяснить результаты индивидуального развития организма онтогенезом, исходя, однако, не только из необходимости той или иной функции в послеродовой период (сосательный рефлекс, дыхательный рефлекс) или из филогенеза (хватательный рефлекс). Те проявляющиеся после рождения рефлексы, которые, по-видимому, объяснимы только как атавизмы или как необходимые средства поддержания жизни, тоже можно считать следствием онтогенетического развития человека. В модели сотворения в большей степени, чем в естественнонаучных выкладках, исходят из того, что отдельные структуры, а также их взаимодействие устроены весьма целесообразно.
Хватательный рефлекс
Во время раннего периода дифференциации зачатки рук выполняют развивающие движения, которые называются хватательным рефлексом роста, так как они являются подготовкой к последующему "хватанию" (рис. 6.31). Сначала происходит рост ручек в стороны и вперед. Затем происходит легкий изгиб в локтях и тем самым рука приближается к передней стенке грудного отдела. После этого ручка подносится ко рту и происходит легкое сжатие кулачка. Типично человеческое расположение пальчиков (противопоставление большого пальца группе из четырех пальцев) можно заметить уже у двадцатимиллиметрового эмбриона (в возрасте 7 недель). Так называемый хватательный рефлекс есть лишь продолжение в послеродовом периоде того, что ребенок уже "пробовал" в предродовом, утробном периоде, а именно хватания во время его роста.
Сосательный рефлекс
Если посмотреть со стороны, то видно, что нижняя и верхняя челюсть образуют друг с другом открытый вперед угол. Когда расходящиеся стороны этого угла удлиняются в процессе роста, то скелет рта эмбриона становится больше, что и приводит к натяжению тканей вокруг него. Как только увеличивается сопротивление натяжению, губы сжимаются и замыкают мягкую часть рта. Но за губами полость рта продолжает увеличиваться. Это приводит к образованию своего рода полости отсасывания. Сосательный рефлекс новорожденного является, таким образом, следствием описанного внутриутробного процесса развития, который похож на процесс сосания.
Дыхательный рефлекс
С увеличением почти круглых по форме сердца и печени у десятимиллиметрового эмбриона увеличивается угол между этими органами, с одной стороны, и позвоночным столбом - с другой. Здесь возникает своего рода свободное пространство, в котором формируются легкие. Этот процесс продолжается с ростом ребер в ходе оформления грудной клетки. Подобно этому позднее в процессе дыхания легкие будут засасывать воздух в увеличивающуюся грудную клетку (рис. 6.32). Итак, в процессе роста эмбрион "подготавливается" к дыхательным движениям в послеродовой период.
Можно говорить о дородовом развитии функций в смысле развития предрасположенности. Утверждение, что "возникновение органа есть уже начало его функции" действительно для всех органов человеческого тела, например, для чувствительных и двигательных нервов. Не существует органов, которые бы не имели функций. Все органы во время своего развития имеют конструктивное значение для процесса роста и подчиняются правилам, которые сегодня широко известны.
Принципы развития
Попытка филогенетического толкования строения человеческого тела - это не естественнонаучный, а исторический метод. Он основывается частично на предположениях, которые в настоящее время опровергаются данными науки. При условии существования живой яйцеклетки (т. е. при условии, когда она принадлежит человеку, может нормально развиваться и реализовывать свои потенции) можно вывести следующие правила и принципы дифференцировки:
- Bo время оплодотворения у человека соединяются гаметы с индивидуально-специфическими хромосомами, присущими человеку. Они не содержат никакой информации, которая могла бы выразить признак, присущий не человеку.
- В ходе развития человека невозможно найти ту грань, которая позволила бы определить момент, до которого человек по своей индивидуально-специфической сути еще не являлся бы Человеком в полном смысле.
- Человеческий организм не является простой суммой частей, но с самого начала своего развития представляет собой единое целое. Из этого следует, что у него не может быть "кирпичиков" из прошлых эпох, а весь организм как таковой, во всех своих свойствах и проявлениях всегда оставался человеческим.
- Каждый орган функционирует в соответствии со свойствами, которыми он на той или иной стадии развития обладает. Каждый шаг в дифференциации следует выводить и объяснять как онтогенетически необходимое качество оплодотворенного яйца.
Поскольку уже было доказано, что каждый орган для процесса роста является конструктивно необходимым, то "функциональная бесполезность" в качестве онтогенетического доказательства филогенетического развития становится несостоятельной. В пользу историко-родового развития говорит еще в некоторой степени двусмысленный аргумент сходства. Приведенный выше пример с хордой особенно наглядно демонстрирует: о функциональной бесполезности не может быть и речи, хотя и имеется сходство с хордой беспозвоночных животных. Различные ранние стадии эмбрионального развития передних конечностей, например, у кита и человека никоим образом не являются функционально бесполезными и походят друг па друга и гораздо большей степени, чем это позволяет предположить окончательный вывод. Но, как было подробно изложено в разделе 6.1, сходство может быть признаком творений, принадлежащих руке одного и того же Творца или указывать на (возможно, неизвестную) конструктивно-морфологическую функциональную необходимость.
Выводы
Из изложенного выше стало понятно, что интерпретация эмбрионального развития человека и других организмов в русле эволюционной теории возможна только как допущение об эволюции, и лишь после этого возможно ее применение при изучении отдельных органов или свойств. Авторам учебника представляется, однако, более соответствующим фактам взгляд на ход событий и положение вещей с точки зрения теории сотворения, который характеризуется следующими положениями.
Так же, как в неорганической природе действует закон о сохранении энергии, так и в живой, органической природе действует закон сохранения индивидуальности. Он гласит, что с оплодотворением возникает индивидуальная специфика организма и его обмена веществ, которая сохраняется до его смерти. То, что меняется в ходе развития, является лишь внешним.
Для человека это означает: человек не становится человеком, но он есть человек с момента зачатия. Он не развивается в человека, но развивается как человек. Человек - это не феномен, который является результатом онтогенеза, это та данность, которая является предпосылкой онтогенеза.
Данные эмбрионального развития человек... имеют значение не только для эволюционного учения, они приобретают особый вес, когда речь идет об убийстве нерожденной жизни. И даже если это не принимается во внимание авторами школьных учебников и учителями биологии, все же невозможно отрицать, что "основное биогенетическое правило" формирует у учеников представление, что человек, особенно на начальных стадиях своего онтогенеза, является как бы "не совсем человеком", а скорее похож на животное. И если учесть, что только в Федеративной Германии ежегодно в материнской утробе убивают около 300 000 детей, то закон о сохранении индивидуальности не может быть недооценен и оставлен вне поля зрения. И тут в полной мере становится ясно, что представление о человеке, находящееся в основе нашего мышления, определяет действия и поступки людей.
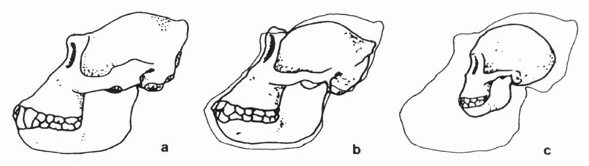
Рис. 6.19. а - череп взрослой мужской особи гориллы; b - череп женской особи гориллы среднего возраста; с: череп молодой, еще не выросшей особи гориллы. Контур черепа особи а был наложен на изображение черепов особей в и с. Череп молодой гориллы имеет гораздо больше сходства с черепом человека, чем. череп взрослой гориллы, однако, вряд ли биологи применят основное биогенетическое правило в этом случае и сделают вывод о происхождении гориллы от "человекоподобного предка . (Sengbusch, 1985, Eckhard.)
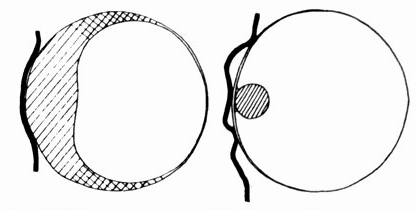
Рис. 6.20. человеческая яйцеклетка (слева) явно отличается от яйцеклетки макаки резус (справа) на стадии прикрепления к слизистой оболочке матки /толстая черная линия/. Переход толстостенной части яйцеклетки в тонкостенную слева постепенный, справа - резкий.

Рис. 6.21. Коллекция документальных данных исследований человеческих эмбрионов Блехшмидта. (Анатомический музей университета г. Геттенген.)
Рис. 6.22. Человеческий эмбрион величиной 3,4 мм в возрасте 27 дней. Голова наклонена над областью сердечного утолщения. При этом на изогнутой стороне возникают характерные складки (область зачатков лица).
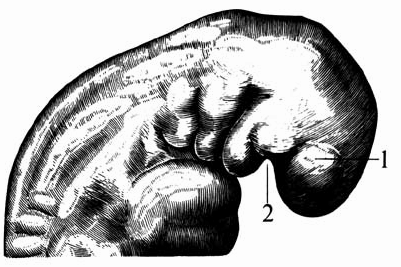
Рис. 6.23. Человеческий эмбрион величиной 3,4 мм. Реконструкция области головы г ясно различимыми складками изгиба (висцеральные дуги). 1 - зачатки глаз: 2 - ротовое отверстие и уплотнение верхней челюсти; под ними - дуга нижней челюсти, подъязычная дуга и первая дуга гортани.
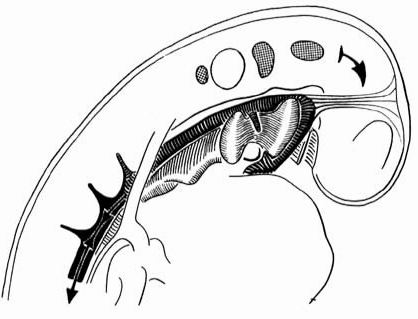
Рис. 6.24. Схематическое изображение изгиба человеческого эмбриона в процессе роста. Зачаток головного мозга растет в длину больше, чем питающий парный кровеносный сосуд (черный цвет на рисунке). Тем самым головной мозг, связанный с сосудами тканями, "сдерживается в росте".
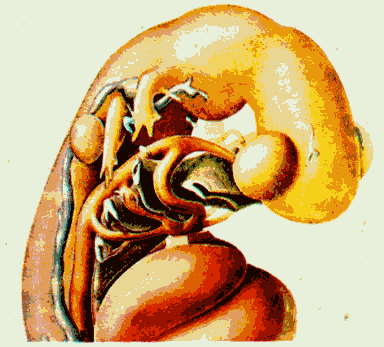
Рис. 6.25. Строение человеческого эмбриона величиной в 3,4 мм в области головы. Сосуды внутренних дуг образуют короткие смычки между брюшной аортой и парной спинной аортой. Они обхватывают головную кишку.
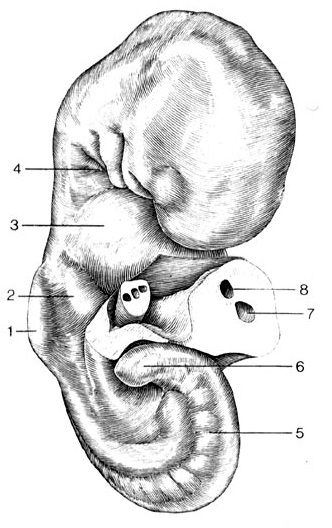
Рис. 6.26. Человеческий эмбрион в возрасте 28 дней, размером в 4,2 мм, в разрезе. 1. Зачаток рука; 2. Утолщение а области печени: 3. Утолщение в области сердца. 4.Складки изгиба; 5. Мета-мерны стенок тела; 6. Нижний конец тела; 7,8. Кровеносные сосуды пуповины.
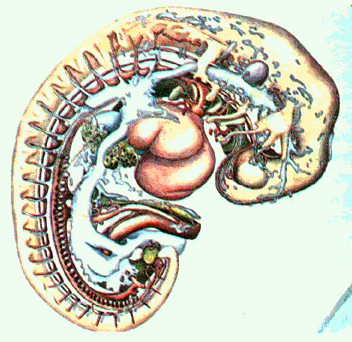
Рис. 6.27. Экспонат, реконструирующий человеческий эмбрион в возрасте 28 дней размером 4,2 мм, в разрезе.
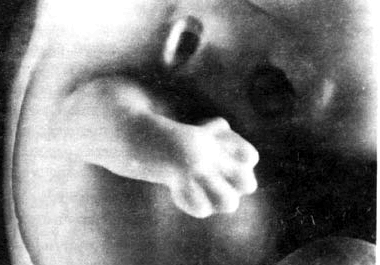
Рис. 6.28. Рука и ладонь человеческого эмбриона в начале седьмой недели развитии величиной 15,5 мм. Пальцы в зачаточном виде представляют собой сначала лишь маленькие бугорки по краю ладони. Но теперь их уже можно отчетливо различить. Они вoвce не образуют "плавательную перепонку".
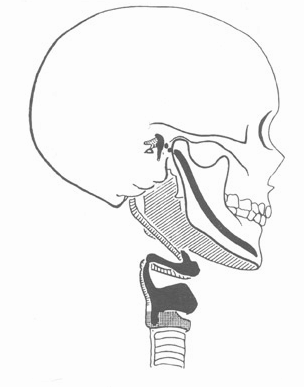
Рис. 6.29. Человеческий череп (взрослого индивидуума). Первичный и вторичный челюстной сустав. Меккелев хрящ в нижней челюсти (обозначен черным цветом) с развитием ротового отверстия (удлинением, скелета верхней и нижней челюсти) опрокидывался" относительно положения, почти параллельного основанию черепа, и при этом подразделялся на фрагменты (точки на рисунке). Из этих фрагментов образовывались слуховые косточки. По мере окостенения хряща и роста скелета нижней челюсти у основания черепа вновь формировался вторичный челюстной скелет. (Dwight из Беннингхоффа, 1968.)
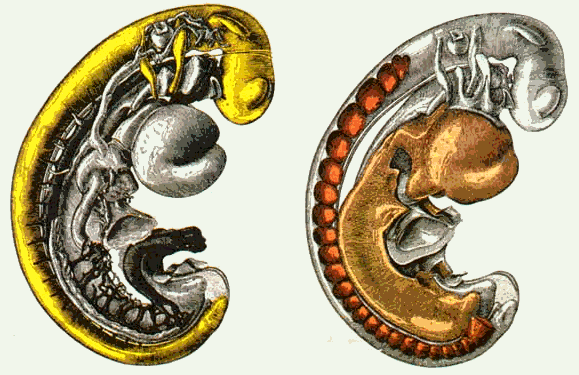
Рис 6.30. Метамерные структуры человеческого эмбриона величиной 2,57 мм в возрасте 26 дней, реконструкция строения сделана по серийным срезам. Слева: желтым показан спинной мозг с питающими его (метамерными) сосудами: справа: темно-красным - метамерные органы тела, возникающие в полостях между сосудами, зачатки мускулов и т. д.
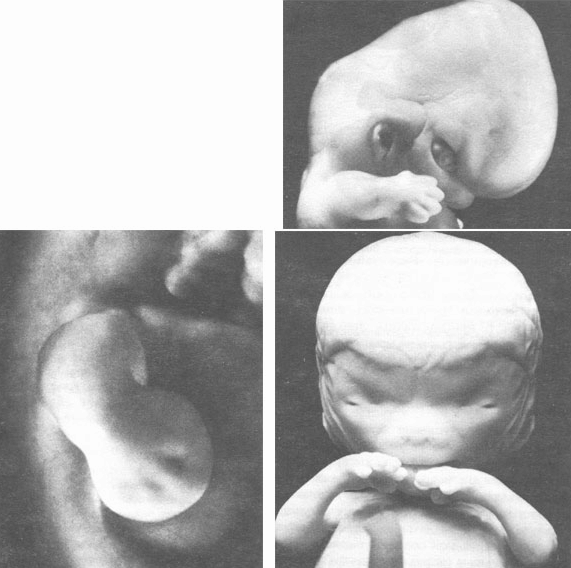
Рис. 6.31. Хватательный рефлекс человеческого эмбриона. Развивающие движения ручек эмбриона являются ранними хватательными движениями со сжатием большого пальца и кулачка. Эмбрионы различной величины: 10,6 мм (слева), 18 мм (сверху) 24 мм (внизу).
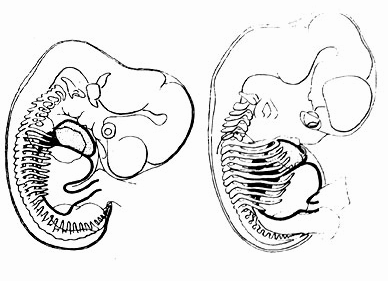
Рис. 6.32. Человеческий эмбрион величиной 10 мм и 17,5 мм. Сердце обозначено пунктиром, печень - жирной черной линией. В то время как сердце и печень, увеличиваются, становится больше и угол между печенью, сердцем и позвоночным столбом. При этом с развитием грудной клетки происходит и развитие легких как органа эмбрионального дыхания.



