




История происхождения и развития жизни
3.3.1. Возможности и границы мутационных процессов
В этом разделе рассматриваются следующие вопросы:
-
К каким научным выводам приходят на основании эмпирических выводов при исследовании мутаций?
-
Может ли суммирование множества мутационных изменений привести к макроэволюционному изменению?
Получить окончательный ответ на второй вопрос можно лишь с учетом других эволюционных факторов (особенно отбора).
В качестве ответа на первый вопрос мы выдвигаем следующее утверждение:
Мутационный процесс - насколько можно констатировать на основании эмпирических фактов - остается в области микроэволюции.
Это утверждение обосновывается и иллюстрируется целым рядом примеров подобных мутаций (см. табл. 3.1; рис. 3.7 - 3.13). Так как у мушки дрозофилы, излюбленного "конька" исследователей мутаций, известно необозримое число мутаций (рис. 3.7), то этот пример можно считать наилучшим и наиболее полным сводом известных на сегодняшний день мутаций, хорошо изученных генетиками. Следует принимать во внимание также мутационные изменения, произошедшие в результате целенаправленной деятельности человека (селекции).
Примеры из таблиц табл. 3.1 (см. также табл. 3.2 и 3.3 в разделе 3.3.5), все без исключений, показывают, что посредством мутаций происходят только изменения уже имеющихся структур. Это касается и селекционно-позитивных мутаций (сравните с этим примеры из раздела 3.3.3). "Изменение" может означать:
-
увеличение количества содержимого вещества (например, содержания сахара в сахарной свекле, пигмента при цветении);
-
деформацию (например, крыловые мутанты Drosophilia);
- увеличение какого-либо органа (например, цветков или плодов);
-
изменение количества частей тела (например, дополнительные ребра у свиньи).
Даже экстремальные случаи, как. например, четырехкрылые мушки (tetraptera -мутанты), подтверждают тот же принцип: удвоение уже имеющейся структуры "пара крыльев" делает их неспособными к полету, так как у них отсутствуют жужжальца. В основе регуляции развития лежат, как в этом примере, многократные изменения и дефекты (мутации регуляторных генов), прежде всего при появлении атипичных структур. При этом атипичную структуру нельзя путать с "новой структурой" (например, формы плакучих растений).
Если бы возникли новые "конструктивные элементы", то можно было бы говорить об истинно эволюционном новообразовании (макроэволюции). Однако до сего времени такие случаи не наблюдались.
Мы можем не касаться вредных мутаций, так как они не могут оказывать длительного воздействия на генофонд вида, они элиминируются селекцией.
Дальнейшие рассуждения о целях и результатах деятельности человека в области выращивания растений и животных вы найдете в разделе 3.3.5.
Итак, мы вплотную подошли к рассмотрению второго вопроса этого раздела: может ли суммирование множества микроэволюционных изменений вызвать микроэволюцию (рис. 3.10)? Результаты проводимых до сих пор мутационных исследований позволяют дать скорее отрицательный ответ, чем положительный. Это особенно наглядно демонстрируют исследования плодовой мушки Drosophilia, насчитывающей более тысячи поколений и миллионы особей: все мутанты Drosophilia являются мухами, относящимися к одному и тому же виду Drosophila. Происходят лишь изменения отдельных органов и признаков, которые уже были в наличии и до того. Мутационный процесс остается в рамках микроэволюции, а в данном случае даже внутри одного вида: существуют близкородственные виды, которые почти не отличаются друг от друга морфологически (но внешнему виду), но в природе репродуктивно изолированы, т. е. не производят гибридов. Это виды-близнецы (рис. 3.51).
Генетик В. Готтшлак приходит к выводу: "Новые виды экспериментально не создаются ни путем постепенной аккумуляции генетических мутаций, ни индуцированием отдельных прогрессивных мутаций" (1978, с. 284). Рремане, Шторх и Велш пишут в "Эволюции" (1980, с. 175):
"Мы знаем... множество случаев, когда у мутантов исчезает дифференцировка: например, горох с листьями без усиков. У цветов, чашечка которых превратилась во второй венчик, исчезла дифференцировка между лепестками и чашелистиками... Но нам не известны истинные мутации генов, ответственных за дифференцировку".
Такие мутации дифференцировки, т.е. мутации, следствием которых является превращение исходной ткани в совершенно отличную от нее дочернюю ткань, как раз и были бы необходимы, для того, чтобы могла совершиться макроэволюция. Рис. 3.14 демонстрирует обратный случай.
Итак, если в ходе мутаций изменяются только уже имеющиеся структуры, то из простого логического рассуждения становится ясно, что даже суммирование множества мутационных изменений не может приводить к образованию новых форм.
Возможно привести следующий аргумент: в прошлом происходило, вероятно, больше положительных мутаций. Но против этого говорит тот факт, что вымершие существа были прекрасно приспособлены к условиям окружающей среды - это неоспоримый факт палеонтологии (см. главу 8).
Нужно исходить из того, что мутации большей частью вредны и названный аргумент поэтому неоснователен (для аргументации следовало бы принимать в расчет длительные временные периоды; см. раздел 3.5.1).
Заключение
Эмпирические результаты мутационных исследований не дают доказательств
макроэволюции. Изученные до настоящего времени факты позволяют сделать вывод
о том, что для мутационных процессов определены достаточно узкие рамки, а выход
изменчивости организмов за рамки микроэволюции обосновать известными мутациями
невозможно.
| Таблица 3.1. Примеры мутации |
| a) В мире животных: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| б) в мире растений: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| в) бактерии: |
|
|
|
|
|
|
|
|
| г) человек: |
|
|
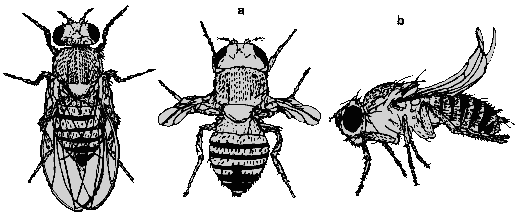
Рис. 3.7. Плодовая мушка дрозофила: исходный тип
( = нормальная форма) слева; а также две разновидности мутантов, а - vistigal
- рудиментарные крылья, в - unturned - загнутые крылья. (По Dremerec., Kaufmann,
1978 ч Lindsley, Grell, 1972.)
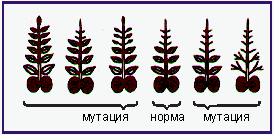
Рис. 3.8. Виды мутации листьев гороха. Листья нормальной формы состоят из трех пар перистых листьев, усиков и прилистника” (обозначены черным). Мутанты имеют большее или меньшее число перистых листиков. В отдельных случаях они имеют либо только перистые листики, либо - только усы. (По Gottschalk, 1978.)

Рис. 3.9. Мутанты вороньего глаза (Paris quadrifolio) с пятью листковыми, чашечковыми и цветковыми листиками, десятью мужскими цветками и с пятигранной завязью (внизу). Цветки нормальной формы (наверху) состоят из четырех лепестков (и тем самым, в общей сложности, из восьми мужских лепестков). Мутация не произвела новой структуры.
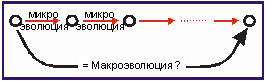
Рис. 3.10. Суммирование множества микроэволюционных шагов и макроэволюция. Может ли макроэволюция наступить в результате множества микроэволюционных шагов?

Рис. 3.11. Альбиносы (здесь: проявление альбинизма у гадючьего лука [Muscari]) появляется вследствие потери организмом способности синтезировать определённый пигмент. Мутанты-альбиносы часто встречаются и в царстве животных, и в царстве растений. Бывают альбиносами и люди.
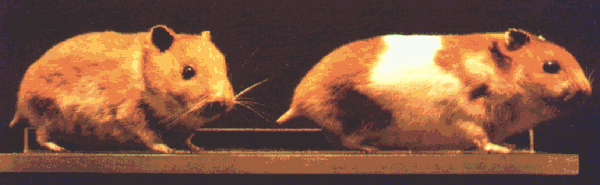
Рис. 3.12. Золотистый хомячок, завезённый в Европу из Сирии в 1930 году, разводился в течение многих лет. В I960 году появился мутант с пятнистой шкуркой (справа). (Вестфальский краеведческий музей, Мюнстер.)

Рис.3.13. Мутанты волнистого попугайчика, потерявшие первоначальную окраску: у левого экземпляра отсутствует желтый цвет в окраске оперения. Справа: нормальная форма. (Вестфальский краеведческий музей, Мюнстер.)

Рис. 3.14. Роза с позеленевшими цветами - пример
дедифференциации: частично ликвидированы различия между лепестками цветка и
чашечки. Обратный процесс дифференциальной мутации пока нигде не наблюдался



