




История происхождения и развития жизни
10.6. Изменчивость внутри основных типов
Из определения основного типа можно сделать вывод о том, что все принадлежащие к единому основному типу (животные, растения и т. д.) обладают одинаковым паттерном последовательности расположения генов,- иначе не было бы возможным никакое скрещивание. Подобное сходство генетического паттерна наглядно проявляется во внешнем виде (фенотипе), поэтому известные на сегодняшний день основные типы совпадают зачастую с известными давно уже таксономическими группами, приблизительно родами или семействами (рис. 10.15), однако не всегда и не обязательно. Обычно для ученых, занимающихся таксономией, свойственно длительное, очень тщательное и точное проведение наблюдений за природой, которые, как правило, на заключительном этапе проходят проверку исследованиями в области скрещивания. Этому же следуют в своих исследованиях и современная молекулярная биология и биохимия. Более тщательному изучению в этом плане было подвергнуто семейство утиных, и при этом выяснилось, что утиные по всем названным выше признакам явно отличаются от всех остальных птиц, исследованных до сих пор. На рис. 10.16 изображен целый ряд птиц из семейства утиных, хотя бы даже по характерной форме утиного клюва. Однако, с другой стороны, сразу же бросается в глаза, насколько эти птицы различны по величине, форме, цвету и даже по образу жизни (см. также рис. 10.2 и 10.4). Согласно модели основного типа, в ней содержится возможность в рамках учения о сотворении вывести от одной (или нескольких) сотворенных форм происхождение всех видов, принадлежащих к единому основному типу, и проследить развитие этих видов от одной формы в ходе микроэволюционных процессов. Лошадь, таким образом, действительно имела "пралошадь" в качестве предка! Однако эта "пралошадь" (и в этом специфика учения о сотворении) обладала, как типичная лошадь, наряду со всеми основными признаками современной лошади, гораздо большим потенциалом изменчивости, чем ее потомки. Подобные положения постулируются и для исходной формы утиных, для куриных, собачьих, для яблоневых и т. д.
Может ли подобная гипотеза быть доказана фактами из жизни природы? Докажем это хотя бы по нескольким пунктам на примере представителей семейства утиных. Мы уже упоминали об этом: некоторые утки не скрещивались с другими. И среди них множество форм, с которыми у таксономов всегда было много проблем. Этой группе присвоили название "аберрантные типы". Они отличаются тем, что объединяют в себе признаки совершенно различных утиных родов. Поэтому они не могут быть причислены к другим группам утиных, а составляют свою собственную ветвь. "Куриный гусь" (рис. 10.17) является примером подобного аберрантного типа и соединяет в себе признаки полугусей, гусей и уток. "Пароходные утки" по строению своих протеинов и перьям ближе к полугусям, по признакам строения скелета - к уткам, а по составу смазки крестцовой железы их можно считать близкими к гаге обыкновенной. Подобные проявления противоречий в признаках встречаются довольно часто и в рамках эволюционной модели могут объясняться лишь конвергенцией (см. раздел 10.7). Согласно этой модели, на первый план выдвигается гипотеза давления отбора, который и привел к появлению в различных видах одинаковых признаков. Однако подобное давление отбора неизвестно. Модель сотворения предполагает, что в исходной форме утиных был заложен большой запас возможностей для выражения и передачи потомкам тех или иных признаков в зависимости от требований условий, времени и большей или меньшей степени случайности комбинаций. Это касается как окраски оперения, так и формы клюва. Морфология гибридов из семейства утиных показывает, что у различных видов обнаруживаются достаточно схожие генетические паттерны, отчетливо проявляющиеся в ходе гибридизации. На рис. 10.18 показано, что гибриды между чернетью хохлатой и нырком красноголовым с малой морской чернетью похожи настолько, что их можно спутать.
Исследования в области поведения тоже дают аргументы в пользу модели основного типа. Конрад лоренц, применяя новаторские методы исследования поведения уток, выяснил, что различные поведенческие процессы расчленяются на отдельные элементарные двигательные акты, которые свойственны почти всем уткам. Согласно гипотезе конрада лоренца, новая схема поведения возникает либо вследствие проявления новой комбинации элементарных двигательных актов, либо вследствие утраты какого-то отдельного элемента. Образование различных схем поведения внутри семейства утиных может, таким образом, трактоваться, вне всякого сомнения, как микроэволюционная вариация "основного образца" утиного поведения, предусматривающего высокую способность к изменчивости.
Мощную поддержку модели основного типа оказывает и тот факт, что ученые обнаружили абсолютно определенные связи между утками даже на уровне биохимического родства. Примером тому может служить протеин лизоцим. Это энзим, который может разрушать стенки бактерий. Среди птиц семейства утиных проявляются две четко различимые формы. Исследовали 25 видов уток, в 5 из них обнаружили первый тип лизоцима, в 3 видах - второй тип лизоцима, а в 13 видах были найдены оба типа лизоцима одновременно. У 4 видов ученые не смогли классифицировать принадлежность лизоцима ни к первому, ни ко второму типу. И вновь обнаруживается, что 4 группы вообще не являются связанными таксономически друг с другом: каждая группа состоит из представителей различных видов. Согласно эволюционно-теоретической модели, подобное распределение могло бы возникнуть в результате конвергенции, однако, необходимое для этого давление отбора остается опять-таки "тайной за семью печатями". В то же время модель основного типа, напротив, дает хорошую возможность трактования этого факта (рис. 10.19). Исходная форма утиных имела, вероятно, оба типа лизоцима (была, т. о., сложной), затем некоторые потомки утратили один, другие второй энзим. У отдельных видов оставшийся лизоцим претерпел изменения вследствие точечных мутаций, при этом по своей функции он остался неизменным, однако причислить его напрямую ни к одному из основных типов не представляется возможным.
Следует отметить, что подобные микроэволюционные процессы могут протекать чрезвычайно быстро: так, например, аминокислотная цепочка d-гемоглобина домашней утки отличается от подобной цепочки у дикой утки двумя позициями, а цепочка р-гемоглобина - даже пятью позициями! Всего лишь за несколько сотен лет, максимум в течение тысячелетия, благодаря искусственной селекции было достигнуто такое явное различие. Однако это не помешало утке остаться уткой, а измененный гемоглобин, кажется, функционирует не хуже и не лучше первоначального, наблюдавшегося у диких форм.
Правило обратной изменчивости.
Наблюдения за различными организмами, проводимые вновь и вновь в течение десятилетий, показали, что спектр различий между мутантами, получаемыми в результате многократного повторения искусственных мутаций, увеличился лишь в незначительной степени. Это означает, что в ходе этого процесса все время появлялись одинаковые мутанты. Генетик готшалк пишет (1978 г., с. 168): "Чем больше разнообразие, тем труднее расширить его за счет новых мутационных типов. При этом все время возникают мутанты, которые уже существуют". Селекционеры пришли к выводу, что в результате искусственных мутаций посредством облучения или обработки химическими веществами возникают в основном свойства и признаки, которые возникают и в результате спонтанных мутаций. Количество новых типов, полученных в ходе мутаций, неуклонно снижается с увеличением числа попыток проведения мутаций. По этим причинам мутационная селекция потеряла свое значение. Феномен частого повторения одних и тех же мутаций aehhmi (1988) формулирует как "правило обратимой изменчивости". Регулярность, с которой проявляются мутации, указывает на заранее заданный, богатый, но, в конечном итоге, ограниченный потенциал изменчивости, что вполне соответствует требованиям модели сотворения.
Концепция основного типа и эволюционная концепция.
Зададим себе вопрос: а не является ли учение о сотворении, в конце концов, ни чем иным, как особым видом эволюционного учения? Если понимать иод эволюцией только процесс микро-эволюции, то есть вариации уже имеющегося без возможности создавать нечто абсолютно повое (макроэволюция), то это верно. Однако, ни один сторонник эволюционной теории не согласился бы с этим. Как уже было детально изложено в этом учебнике (разделы 3.3 и 4.4), эволюционные механизмы, поддающиеся экспериментальной проверке, могут только лишь изменять имеющуюся в наличии информацию, однако, на современном уровне науки уже доказано, что ничего принципиально нового они произвести не могут. В этом модель сотворения и эволюционная модель в корне расходятся. Внутри основною типа это может привести к расщеплению на различные биологические виды, которые затем могут заселять различные экологические ниши - как, например, нырки в противовес водоплавающим уткам и крохалям - и при этом остаться вблизи от утиных. Однако концепция основного типа не предусматривает, что из основного типа утиных, куриных, лошадей, собак или человека с помощью механизма случайностей не могут возникать другие основные типы и что наблюдаемые в сегодняшнем мире общности также не произошли от других основных типов.
Как эволюционная модель, так и модель сотворения постулируют исходные формы, однако, ни в одной из двух моделей до сих пор наличие их не может быть доказано. В рамках модели сотворения изначальные формы считаются, в противоположность эволюционной модели, не более "примитивными", а более "сложными", чем их современные потомки.
Понять все это может помочь одно сравнение. В разделе 6.5 речь шла об эмбриональном развитии человека. Из одной яйцеклетки в течение всего лишь нескольких месяцев развивается множество тканей и органов с различными типами клеток. Этот процесс называется дифференциацией. Клетки, прошедшие дифференциацию, существенно отличаются от яйцеклетки: они не могут служить исходным пунктом образования нового организма, потенциал их развития становится меньше в процессе дифференциации.
В рамках модели сотворения подобный смысл вкладывается и в понятия "развитие" или "микроэволюция". Потенциал развития оплодотворенной яйцеклетки равен гипотетическому потенциалу изменчивости сотворенной исходной формы. Как во время эмбрионального развития информационное содержание организма не возрастает, хотя при этом возрастает многообразие клеточных и тканевых типов, так и не становится большей сложность биологической информации в ходе постулируемой дифференциации основных типов в биологические виды. Потенциал изменчивости отдельных видов со временем снижаются, тогда как степень специализации постоянно возрастает (см. раздел 3.4 о принципах образования вида, особенно рис. 3.57-3.50). Уместно будет в данном случае вспомнить еще раз о механизмах изменчивости (см. разделы 4.2 и 4.4).
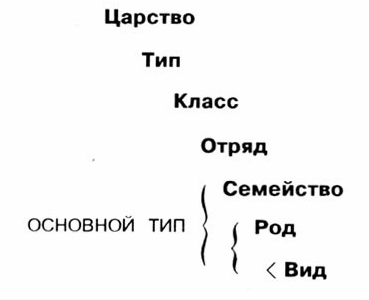
Рис. 10.15. Основные типы могут совпадать с семействами, родами или видами.

Рис. 10.16. На этом, рисунке изображена небольшая часть из множества представителей семейства утиных. Первый ряд: лебедь черношейный (Cygnus melanocoryphus), лебедь-трубач (С. buccinator, самая большая в мире птица из семейства утиных), канадская казарка (Branta canadensis). Второй ряд: африканский малый гусь (Nettapus auritus), лайсанская кряква (Anas platyrhynchos, подвид кряквы), обыкновенная кряква (Anas platyrhynchos laysanensis), краснозобая казарка (Branta ruficollis). Третий ряд: мандаринка (Aix galericulata), длинноносый крохаль (Мегди.ч serrator). Все виды относятся к основному типу утиных. (Рисунок thomas hilsberg.)

Рис. 10.17. "Куриный гусь" представляет собой немалую проблему для ученых-таксономов. Он объединяет в себе признаки абсолютно разных групп утиных. Поэтому его причисляют к "аберрантному типу", который находит убедительные объяснения в рамках модели сотворения.

Рис. 10.18. Изображенная наверху утка - чернеть хохлатая - может скрещиваться с нарисованным под ней красноголовым нырком, в результате чего появляется нарисованный третьим сверху гибрид. R парковых прудах его постоянно путают с проживающей только в Америке малой морской чернетью (изображена внизу). Известно множество подобных случаев, Koiya в результате скрещивания двух видов гибридов проявляется сходство с неким третьим видом. Из приведенного выше примера можно сделать вывод о том, что наследственность задействованных видов является очень схожей и что гены в спокойном состоянии не выражены, но в процессе скрещивания между видами активизируются. (По scherer и hilsberg, 1982.)
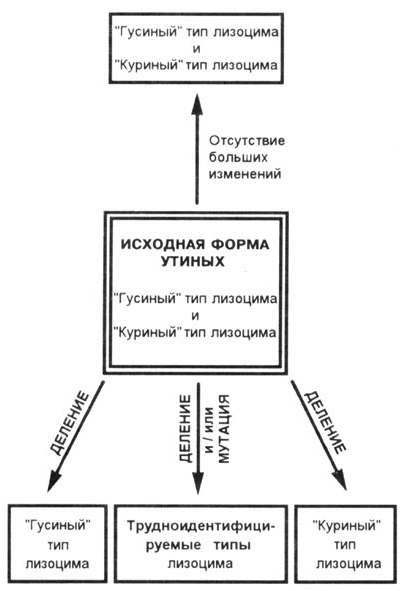
Рис. 10.19. Рисунок представляет нам вариант графического
изображения гипоотезы распределения различных типов лизоцима у птиц семейства
утиных как результат производного из сложной родовой формы. Родовая форма, должно
быть, имела оба типа энзима лизоцима (в названии лизоцим заложены исторические
корни). Следствие делений и различных точечных мутации при содействии механизма
случайности могли возникнуть современные комбинации этою протеина. Подобный
процесс подпадает под область воздействия микроэволюции.



