




История происхождения и развития жизни
10.4. Что такое "основной тип"?
Еще в первых разделах учебника понятие "основной тип" появлялось время от времени, и теперь настало время дать ему определение. О понятии вида в современной биологии подробно говорилось уже в разделе 3.4, оно приводится в таблице 10.3 вместе с другими определениями видов и типов. Там приведена лишь малая часть из всего богатого набора определений, и никого не удивляет тот факт, что в университетских библиотеках можно найти такие книги, как "Философские проблемы теории вида".
Все приведенные в таблице 10.3 принципы классификации объединяет следующее: они не дают четких объективных критериев, которые были бы способны однозначно разграничить организмы. Именно самые уязвимые места трактовки видов с точки зрения биологии, согласно взглядам эрнста майра, изложены в разделе 3.4. Американский ученый, профессор биологии франк Л. марш, в настоящее время оставивший занятия наукой, первым более 40 лет назад предложил определение вида, согласно учению о сотворении, на основании которого построено приводимое ниже определение основного типа: "Все индивидуумы, которые прямо или косвенно связаны путем, скрещивания, либо чьи зародышевые клетки после естественного оплодотворения, по крайней мере, начинают процесс эмбрионального развития с проявлением наследственных черт обоих родителей, причисляются к одному основному типу".
На рис. 10.2 этому определению дается наглядное пояснение. Если, например, обыкновенный фазан скрещивается с пестрым (китайским) фазаном и с домашним петухом, но домашний петух не скрещивается с пестрым фазаном, то они оба не напрямую, но все же связаны посредством обыкновенного фазана и, таким образом, относятся все-таки к одному основному типу. Это же является правомерным и для индейки и пестрого фазана, хотя при этом и не говорится о том, что пока еще не наблюдавшееся скрещивание абсолютно невозможно. Это должно было бы решиться в ходе эксперимента. В определении, кроме упоминания об эмбриональном развитии, речь еще идет о наследовании признаков обоих родителей, потому что партеногенез должен быть исключен. Можно, например, при помощи сперматозоидов направить эмбриональное развитие некоторых яиц с самого начала по пути развития абсолютно другой группы животных, однако наследственные признаки в сперматозоидах будут при этом уничтожены. В некоторых случаях подобные эксперименты удаются при помощи укола шприцем! Скрещивание между обыкновенным фазаном и китайским фазаном дает иногда потомство, способное к воспроизведению, в то время как исследованные до настоящего времени результаты скрещивания между обыкновенным фазаном и домашней курицей не способны к произведению потомства. И, напротив, оплодотворенное индюком яйцо обыкновенного фазана лишь в редких случаях доходит до стадии взрослого гибрида, в то время как произведенное искусственным путем скрещивание между домашней курицей и индейкой дает отрицательный результат уже на эмбриональной стадии развития, хотя эмбриональное развитие все же начинается. (При определенных условиях иногда встречается естественное>скрещивание между этими двумя видами к природе). Подобные "скрещивания" принимаются во внимание в приведенном нами выше определении основных типов (в отличие от определения биологического вида). На рис. 10.2 изображена дополнительно кряква, о скрещиваниях которой с какими-либо представителями куриных неизвестно, однако известны многие случаи скрещивания с другими утками. Таким образом, кряква не принадлежит к тому же основному типу, к которому относятся четыре перечисленные выше куриные птицы.
Эмбриональное развитие амфибий и рыб более доступно для экспериментов. Скрещиванию подвергались, к примеру, различные географические расы североамериканских леопардовых лягушек (Rana pipiens) между собой. При этом было обнаружено, что эмбриональное развитие гибридов прерывалось все раньше, и связано это было со степенью возрастания географической изоляции рас. Эмбриональное развитие прерывалось тем раньше, чем выше была степень географической изоляции, так как начинали проявляться всяческие аномалии. Отдельные породы так похожи между собой, что классическая таксономия не хочет видеть в них разные виды.
Все сказанное выше в достаточной степени проясняет тот факт, что способность производить потомство, конечно же, не может привлекаться в качестве критерия, характеризующего принадлежность родителей к основному типу.
Стерильность может быть воспроизведена тоже в ходе экспериментов (предположительно, посредством нескольких мутаций в регуляторных областях генов). При :"том последовательность гена остается неизменной (а тем самым также и внешний вид живого существа). Многочисленные поучительные примеры имеются среди множества видов плодовой мушки, два из которых внешне в живом состоянии практически неразличимы, не могут скрещиваться. Итак, если стерильность может быть произведена благодаря сравнительно небольшим изменениям, то она не может быть использована в качестве классификационного признака. И, напротив, плодовитость или начало эмбрионального развития свидетельствует о том, что основа структуры генов задействованных видов идентична.
Что касается молекулярно-биологического уровня, то эта область в определении основного типа пока еще остается открытой. Так, например, можно было бы задаться вопросом, что именно происходит на уровне ДНК, когда ведут речь о выражении наследственных признаков обоих родителей. И, в конце концов, на повестку дня в этом месте наших рассуждений неизбежно всплывает вопрос, на который пока еще нет ответа, какие гены или комбинации генов являются специфическими для того или иного основного типа. Это представляет собой целую область в учении о сотворении, в которой открывается широкое поле для исследования основ биологии.
В самом строгом смысле определение основного типа касается лишь индивидуумов, так как только на этом уровне может быть проверено экспериментально. Из практических соображений мы попытались применить это определение и к биологическим видам. Эго нам удалось без особых трудностей, так как индивидуумы определенного биологического вида, по определению, могут скрещиваться друг с другом.
В следующих разделах мы обсудим разграничение, биологию и микроэволюцию основных типов на основе некоторых примеров. Для нас представляется особенно важным, что теория происхождения будет нами изложена на конкретных примерах, что даст нам возможность услышать в ответ и более конкретную критику.
Таблица 10.3: Перечень некоторых наиболее важных улм классификации живых существ определений. Следует обратить внимание, что все определения достаточно условны ("не слишком жестко" сформулированы), под этим подразумевается то, что они содержат в себе субъективную оценку систематиков.
Биовиды: К биовидам относятся все индивидуумы, которые входят в смешанные естественным образом популяции. Эти популяции изолированы от других подобных групп.
Морфовиды: К морфовиду относятся все индивидуумы, которые совпадают по своим существенным морфологическим, физиологическим и биохимическим признакам.
Круг рас: К кругу рас относятся все географические, замещающие друг друга формы, поскольку в них проявляются переходы, либо от них можно ожидать появления продуктивных гибридов (помесей).
Круг видов: Если объединенные в круг рас формы слишком различны, то перед нами - круг видов (называемый также "супервидом").
Основной план: Основной план - это совокупность строения и функций, свойственная всем без исключения представителям последнего общего предка какой-либо рассматриваемой в данный момент монофилетической группы. Он включает в себя голоморф вида, представляющего данный основной план. Голоморфом называется общее проявление организма в совокупности всех его функций, строения, поведения и онтогенеза.
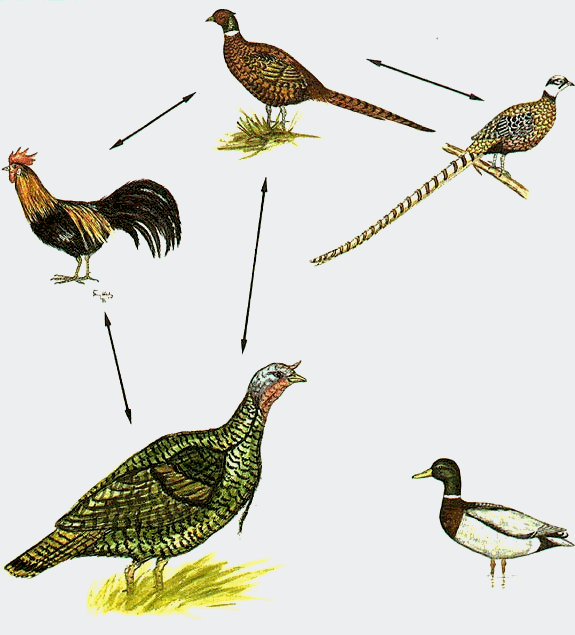
Рис. 10.2. Четыре вида из семейства фазанов (Phasianidae), связанные друг с другом, рядом, скрещиваний. Обыкновенный фазан (Phasianus colchicus) относится к семейству (подсемейству) фазанов, так же как и пестрый китайский фазан (Syrmaticus reevesii) и домашний петух (Callus domestic.us). В то время как индейку (Meleagris gallopavo) причисляют к подсемейству индейковых (Meleagridinae), кряква (Anas platyrhynchos) относится совсем к другому отряду, гусеобразных (Anseriformes). (Рисунок thomas hilsberg.)
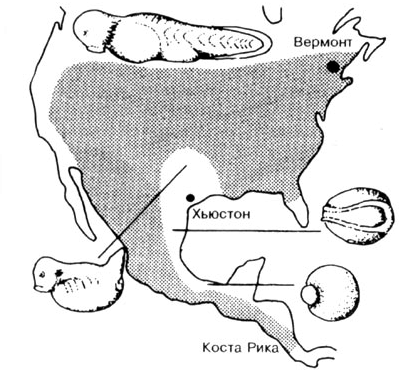
Рис. 10.3. Различные расы леопардовых лягушек (Кала pipiens)
из Северной и Центральной Америки. R ходе экспериментов оплодотворяли яйца географической
расы из Вермонта спермой различных других рас. При этом выяснилось, что в некоторых
случаях скрещивания эмбриональное развитие прекращается, не давая возможности
возникнуть жизнеспособной лягушке. Очевидно, наследственные признаки отдельных
рас благодаря микроэволюционным процессам настолько отдалилась друг от друга,
что гены более не могут быть скоординировано выражены. Интересно, что при этом
важно не лео/рафи-ческое расстояние, а масштаб изоляции в процессе размножения.
Так, например, встречаются скрещивания между расами из Вермонта и Коста-Рики
(расстояние приблизительно равно 6000 км), которые дают жизнеспособные гибриды.
И, напротив, гибриды, получающиеся в результате скрещивания между гибридами
из Вермонта и Хьюстона (расстояние равно приблизительно 2400 км), погибают во
время эмбрионального развития. (По Hadorn и Wehner, 1986, по moore.)



