




Жизнь растений
Экология бактерий
Микроорганизмы распространены повсеместно. Весь земной шар «укутан» в живую пленку, большая доля в которой приходится на микробы. Нет места на нашей планете, где бы не было микроорганизмов. Исключения составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни сверхнизкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни резкие колебания кислотности среды, ни многое другое не мешают существованию и развитию микрофлоры в природных субстратах, правда, в каждом случае разной по составу. Все живые существа — растения, животные и люди — постоянно взаимодействуют с микробами, являясь часто не только их хранилищами, но и распространителями. Горные породы, вода, иловые осадки и почвы также довольно плотно заселены микроорганизмами. Иначе говоря, микроорганизмы — это типичные обитатели — аборигены нашей планеты. Более того, они являются ее первопоселенцами, активно осваивающими самые неподатливые природные субстраты.
Расселением, изучением структуры и функций особей и сообществ организмов в природной обстановке занята специальная отрасль биологии — экология. Исследование микробного мира находится в сфере экологии микроорганизмов. Основная суть этой науки улавливается даже из самого термина «экология» (от греч. «ойкос» — дом, местообитание). Поэтому экологические исследования микроорганизмов проводятся в «их доме». Ни на каком ином уровне, изучая наследственные свойства микробов, химический состав и тонкое строение клетки, нельзя получить тех сведений, которые добываются при экологических исследованиях.
Среды обитания микроорганизмов
Органы и ткани высших организмов
Одной из природных сред обитания микробов являются организмы животных и человека. Прежде всего микробы заселяют кожные покровы, составляя нормальную микрофлору кожи. Кишечник человека также заселен микроорганизмами, не вызывающими заболеваний. Особенно благотворную роль играют молочнокислые бактерии. Мирные взаимоотношения часто нарушаются. Неопасный для человека (и животных) эпибионт (проживающий на поверхности другого организма) проникает в ткани (через раны, царапины), вызывая нагноения. Это свойственно многим неспороносным бактериям, в том числе псевдомонадам.
Совершенно иным субстратом оказывается больной организм человека и животных. Некоторые или многие защитные механизмы и барьеры нарушены, и ослабленный организм становится подобием питательной среды, где развиваются патогенные микробы. Они поражают ткани и органы человека и животных. Даже краткий перечень болезней, вызываемых микробами, заставит любого содрогнуться. Очень мелкие бактерии — риккетсии, внутриклеточные паразиты — возбудители сыпного тифа. В кровяном русле развиваются пастереллы — возбудители чумы. Холера вызывается вибрионом, поселяющимся в кишечнике. Туда же попадают и развиваются сальмонеллы, приводящие к развитию тяжелых заболеваний типа брюшного тифа. Эпидемический цереброспинальный менингит, опасный своими осложнениями, вызывается мелкими кокками из рода Neisseria — организмами в высшей степени адаптированными к паразитическому образу жизни. Многие кокки являются возбудителями пневмонии, вызывают повреждение клапанов сердца. Дифтерия (коринебактерии), туберкулез и проказа (микобактерии) и многие другие болезни вызваны развитием микроба-возбудителя в среде его обитания — в клетках, тканях и органах человеческого и животного (многие теплокровные животные болеют сходными болезнями) организмов. Тяжелейшие болезни вызываются спорообразующими бактериями, среди них газовая гангрена (Gl. perfringens), столбняк (Gl. tetani), сибирская язва (Вас. anthracis) и др.
Атака микробов — возбудителей болезней на человеческий или животный организм проходит не всегда успешно и требует завоевания микроорганизмами их среды обитания (многие облигатные паразиты не могут существовать в иных условиях). Организмы и их органы активно защищаются от инфекции. Барьерами, препятствующими колонизации тканей высших организмов, оказываются разные вещества и структуры:
1) кожа защищается от поселения микробов жирными кислотами;
2) слизистая оболочка носа и глаз — лизоцимом (ферментом, разрушающим клеточные стенки бактерий);
3) кровь — фагоцитами и антителами;
4) ткани рыб — протаминами;
5) корни растений — корковым слоем;
6) фрукты — кутикулой и кислотами;
7) деревья — смолами, таннином;
8) ткани растений — фенольными соединениями, гликозидами.
К такой ситуации оказывается приспособленным и микробный мир. Микробы поселяются в организмах промежуточных хозяев. Часто хозяевами являются многие насекомые, нематоды (черви), животные (особенно опасны грызуны), птицы и даже человек (бацилло- и ви-русоносители, оставаясь здоровыми, опасны для окружающих). Промежуточные хозяева составляют резервуар (очаг) заболеваний, из которого часто развиваются эпидемии. Например, промежуточными хозяевами, составляющими очаг сибирской язвы, являются свиньи, овцы, крупный рогатый скот. Резервуары чумы связаны с грызунами. Переносчиками микробов — возбудителей болезней наиболее часто бывают членистоногие (клещи, вши, блохи). Сходные заболевания (например, лептоспирозы) передаются через разных хозяев (крысы, собаки, лошади). Есть и другие пути передачи инфекции: ветром и насекомыми на сотни километров, при акклиматизации животных в новых районах и т. п.
То, что характерно для человека и животных, свойственно и растениям. Здесь также имеются мирные сожители — эпифиты — обитатели поверхности организмов. Болезнетворные микробы (фитопатогенные) вызывают заболевания растений, часто массовые (подобие эпидемий).
Исключительный случай взаимоотношения микробов с другими организмами представляют собой многочисленные примеры симбиоза (взаимополезного сожительства) микроорганизмов и высших организмов, стоящих на разных уровнях организации. Иногда даже трудно определить, является высший организм субстратом и микробы размножаются в нем либо, наоборот, высшие организмы паразитируют на микробах. Так, например, клубеньковые бактерии образуют на корнях (чаще бобовых) растений наросты, заселенные бактериями (растение — среда для бактерий). С течением времени бактерии разрушаются в клубеньках и растение использует вещества, запасенные микробами (паразитизм растения на бактериях). Подобных примеров можно привести много. В пищеварительном тракте насекомых (лучше изучены муравьи и термиты) имеется свой неповторимый ценоз (ассоциация микробных видов), так как эта крошечная «лаборатория» обеспечивает существование замкнутого микробного мирка в специфических условиях. Нормальное развитие некоторых высших организмов невозможно без содружества с микробами. Известно, что жучки-точильщики (разрушают мебель, древесину) способны питаться древесиной, благодаря тому что целлюлоза перерабатывается в кишечнике их сожителями — бактериями. Питание жвачных животных теснейшим образом связано с активной деятельностью сообщества строгих анаэробов — бактерий в рубце (отделе желудка), где они участвуют в переработке растительных кормов. Клещи, питающиеся кровью высших животных, имеют в своем теле специальный орган, переполненный симбиотическими бактериями, призванными переваривать кровь. Подобными органами обладают все сосущие соки растений насекомые (цикадки). Они имеют в своем теле до пяти разных симбиотических микробов. Удаление симбиотических бактерий может привести к гибели высший организм, так как нарушается обеспечение разными источниками пищи. Так, например, обыкновенный желтый таракан имеет симбиотические дрожжи, которые помогают ему усваивать минеральную серу. Как правило, симбиоз основывается на различиях в обмене веществ обоих компонентов.
Абиогенные субстраты
Главными средами обитания и резервуарами микроорганизмов в природе являются прежде всего почвы, придонные и прибрежные илы водоемов, вода рек, озер и океанов.
Наименьшая плотность заселения микробами характерна для океанических вод. Сюда со стоком рек постоянно вливаются струи, обогащенные микробами и органическим веществом, которые переносятся и распыляются течениями. Мировой океан, занимающий большую часть поверхности планеты, является собирателем и хранителем огромной микробной биомассы. Океанская толща воды играет роль скорее транспортного субстрата, места переживания, чем места активной деятельности микробов. Примером типичного транспортного субстрата может быть воздух. В СССР большие работы в области морской микробиологии проведены А. Е. Криссом.
Вода мелких пресных водоемов (озер) имеет слоистую структуру. Для озер характерны сезонные и периодические процессы. Большое значение для формирования микробных ассоциаций имеют придонные иловые отложения. В то же время прибрежные зоны сходны с почвой. Содержание микроорганизмов в воде пресных водоемов ниже, чем в почвах и илах, но достаточно высоко (1,4 млн. клеток в 1 см3 воды и более). При анализе качественного состава микрофлоры обнаружены представители всех основных физиологических групп микроорганизмов, обеспечивающих круговорот азота, углерода, фосфора и других элементов. В озерной воде отмечается четкое вертикальное распределение бактерий. Максимальное количество бактерий отмечается в летний период, причем в некоторых озерах на значительной глубине (10 м), где было наибольшее количество отмерших водорослей.
Резко отличную картину, характеризующуюся более высокой стабильностью, можно наблюдать в донных иловых отложениях. Главный отличительный их признак — микрослоистость сложения. Это явление связано с тем, что в результате сезонных процессов распределение микроорганизмов происходит неравномерно. Формирование ила при участии микробов идет в тончайших слоях, измеряемых долями миллиметра. В толще, равной всего 1—2 мм, можно обнаружить 6—8 слоев, различающихся составом микрофлоры и физико-химическими условиями. Среди микробного населения илов преимущественное развитие имеют гетеротрофные организмы, аккумулирующие железо и марганец; микоплазмы, стебельковые и почкующиеся бактерии. Ценнейшие исследования по микрофлоре воды и илов проведены в нашей стране широко известными учеными Б. В. Перфильевым и С. Н. Кузнецовым.
Субстратами, резко отличными от илов, где слои обитания микробов по крупицам «лепятся» осадками из толщи воды, являются горные породы, частично выветренные (рухляки), породы из шахт, выброшенные при добыче руд, сами стенки штолен и естественные разломы земной коры. Все это — своеобразная среда обитания микрофлоры, арена деятельности преимущественно хемотрофных и миксотроф-ных бактерий, добывающих энергию при окислении неорганических веществ (соединений железа, марганца, серы). Содержание сапрофитных бактерий в горных породах невелико, около 2—3 тыс. клеток в 1 г субстрата. В нефтеносных породах даже на глубине 500 и 700 м насчитывается 30—100 млн. клеток в 1 г породы. Глубинные воды практически лишены микроорганизмов (3—300 клеток). Микрофлора горных пород активно участвует в их выветривании (разрушении) за счет продуктов жизнедеятельности, в том числе серной и других кислот. Легко догадаться, что такие вещества, как торф, бурый и каменный уголь, содержащие лигнин и гумусовые кислоты, также являются пригодными для микробов субстратами. Нефть и углеводороды успешно осваиваются микроорганизмами. Именно с этими процессами связаны надежды (и уже реальные успехи) современного человечества на получение промышленным путем микробной массы (белка) из нефтяных продуктов для использования в качестве корма и пищи. Микробы способны окислять сульфидные руды, выщелачивая серу и повышая содержание металла в них (в первую очередь цветных и редких металлов). Эти процессы лежат в основе активно развивающейся отрасли промышленности — гидрометаллургии, осваивающей с помощью микробов бедные и трудноперерабатываемые руды.
Почва как среда обитания микробов
Особое место среди природных сред обитания микроорганизмов занимает почва. Это чрезвычайно гетерогенный (разнородный) по структуре субстрат, имеющий микромозаичное строение. Почва представляет собой совокупность множества очень мелких (от долей миллиметра до 3—5 мм) агрегатов (шероховатых гранул, иногда объединенных в группы), пронизанных порами, омываемых почвенным раствором, протекающим по капиллярам.

Рис. 119. Микроструктура почвы. Видны гранулы и очаги. Фото в световом микроскопе 1 — увел. х 20; 2 — увел. х 60.
Эта структура хорошо видна в световом микроскопе (рис. 119); «микропещеры» и туннели-капилляры — могут быть рассмотрены в сканирующем электронном микроскопе (рис. 120).
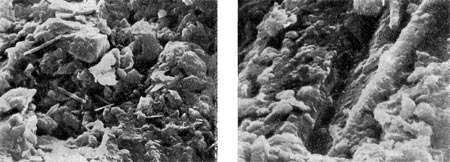
Рис. 120. Микроструктура микробных очагов (фото в сканирующем электронном микроскопе). Увел. X 1600.
Остатки растений и животных, гумусовые вещества — органический элемент почвы — распределены в ней не равномерно, а сосредоточены в отдельных микроочагах, часто устилая пленкой почвенные гранулы, создавая зоны, где протекает бурная, но непродолжительная деятельность микробных сообществ, заселяющих эти участки.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20—100 клеток в каждой). Часто они развиваются в толще сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Природные микроколонии микробов представляют собой размножающиеся в естественной среде популяции, т. е. совокупности родственных между собой особей (клеток) одного вида, расположенных на ограниченном участке природного субстрата. Их развитие начинается с формирования небольших групп клеток. Эти агрегаты клеток (преимущественно бактерий) не бывают хаотическими. Часто они соединены в розетки, спирали и иные организованные группы, образуя первичные микроколонии. Со временем эти первичные микроколонии, расположенные рядом либо разрастающиеся и образующие новые микроскопления, приводят к образованию популяционных колоний, заселяющих в отдельных местах почвенные гранулы (рис. 121).
Диаметр гранул составляет 300—500 мкм. Совокупность гранул образует сложную микроструктуру, состоящую из пор и капилляров, заполненных газами (состав их может быть разным) и почвенным раствором. Такой агрегат является простейшим природным очагом обитания микробов — своеобразной «микропла-неткой». Диаметры подобных «планеток» 3 — 5 мм и более. Если в очаге находится источник энергии (доступное для микробов органическое вещество), то такая совокупность (система) может служить примером простейшей микробной ценотической ячейки — устойчивого в течение определенного времени сообщества микроорганизмов. Главным моментом является присутствие энергетического очага, что создает начало пищевой цепи и служит своеобразным «мотором» для деятельности всего сообщества.
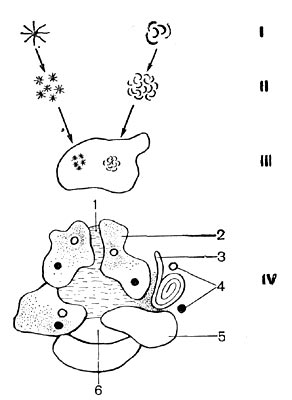
Рис. 121. Схема структуры простейшего микробного ценоза: I — простейшая микроколония (микроагрегат 5—10 мкм), II — популяционная микроколония бактерий (30—50 мкм); III — гранула почвы, заселенная популяционными колониями (300—500 мкм); IV — микроочаг (общий диаметр 0,5—5 лип): 1 — капилляр, заполненный почвенным раствором; 2 — пленка гумусового вещества; 3 — растительный материал; 4 — популяционные колонии микробов; 5 — почвенная минеральная гранула; 6 - пора.
В микроучастке, где находится органический материал (концентрат энергии), поселяются микробы, требующие для своего развития высокой концентрации органического вещества, быстро размножающиеся и минерализующие его. Они являются характерными представителями экологической группы макрофитов — требовательных к пище сапрофитов. Сюда относятся грибы, многие спорообразующие и неспорообразующие бактерии и актиномицеты.
После исчерпания источников пищи деятельность этой группы гетеротрофов в очаге замирает и микробы переходят в состояние длительного покоя, близкого к анабиозу, вплоть до нового притока энергии и новой бурной вспышки их активности. Значительная часть биомассы бурно развившихся популяций микроорганизмов лизируется. Арена активной жизни — гранулы почвы — оказываются усеянными спорами, цистами и другими переживающими формами.
В период высокой активности от очага, обогащенного органическим материалом, в окружающие микроучастки диффундируют растворимые органические вещества (в том числе микробные метаболиты). Это ведет к возникновению зоны, содержащей небольшие (часто следовые) количества легко мобилизуемых источников энергии. В этой сфере развивается микрофлора, растущая при низком содержании пищи в среде. Эти организмы представляют другую экологическую группу микрофитов — сапрофитных микробов, растущих при минимальных концентрациях органических веществ в почве и экономно использующих их. Сюда относятся многие неспорообразующие бактерии, в том числе стебельковые и почкующиеся бактерии, часто известные под названием оли-готрофов (организмов с умеренными потребностями в пище). На богатых (лабораторных) средах они не растут и часто даже погибают.
При новом поступлении органического вещества в очаг тормозится развитие олиготроф-ных бактерий и нередко следует за этим лизис клеток. Часто популяции выживают. Завоеванная олиготрофами территория утрачивается ими. В обогащенном источниками пищи очаге вновь активно размножаются требующие высокого уровня питания сапрофиты. Так возникают микропулъсации активности микроорганизмов и совершается длящаяся миллионы лет смена микробных сообществ. Исследование этих явлений наряду со многими другими: скоростью размножения микроорганизмов, временем индивидуальной жизни и циклами развития в природных условиях — представляют собой сложнейшие проблемы современной экологии микроорганизмов.
Указанные процессы состоят из фаз различной длительности. Одни фазы длятся дольше (разложение значительных масс органического вещества), имеют сезонный характер (растительный опад осенью). Рядом соседствующие микроочаги могут развиваться в противоположных направлениях (обеднение и обогащение очага), что ведет к проявлению упоминавшейся ранее мозаичности. Поэтому почва всегда оказывается насыщенной разного рода микроорганизмами (находящимися, как правило, в состоянии покоя), готовыми к ответу (росту, использованию субстрата) при попадании в почву источников пищи и энергии. Исследования последних лет, проведенные с применением прямых методов учета микроорганизмов в световом и электронном микроскопах, позволили уточнить количество микроорганизмов в почвах и рассчитать их биомассу. Как оказалось, в 1 см3 сравнительно небогатой дерново-подзолистой почвы может содержаться до 20 млрд. клеток. Это составляет около 1% от веса самой почвы. В 30-сантиметровом слое на площади 1 га в почвах разных типов содержится от 1,5 — 2 до 15—40 т биомассы. Живой массой микробов с площади 1 га можно загрузить целую колонну большегрузных автомашин.
По имеющимся в распоряжении ученых данным, биомасса всех обитающих на суше нашей планеты живых существ составляет от 10~12 до 10~13 т (поверхность суши равна 51-Ю9 га), в том числе:
1) суммарная биомасса животных организмов (сухой остаток) — 0,55-109 т;
2) суммарная биомасса растений (сухой остаток) — 55-109 т;
3) суммарная биомасса микроорганизмов (сухой остаток);
а) тундровые и пустынные почвы — 3,06-109т;
б) другие почвы — 71,4-109т;
74,46x1094) биомасса простейших животных и почвенных водорослей (сухой остаток) — 1,5*109 т.
Как легко убедиться, суммарная биомасса животных и высших растений, обитающих на суше, составляет около 56-109 т. В то же время суммарная биомасса микроорганизмов, простейших и водорослей равна 76-109 т. Несмотря на ориентировочность таких расчетов, все же становится очевидным, что биомасса существ, составляющих мир микробов, равна биомассе всех остальных обитателей суши и, возможно, превышает ее. Читатель может сделать некоторые выводы об огромном влиянии, оказываемом на природную среду микроорганизмами.
Сообщества любых организмов, в том числе микроорганизмов, представляют собой не случайные скопления, а организованные объединения популяций, обладающих коллективными функциями и взаимодействием. Такие объединения обладают большой стабильностью, и многие воздействия на среду обитания (вспашка почв, внесение удобрений и др.) не разрушают типичных для данного субстрата ценозов. Сообщества и популяции, их составляющие, обладают рядом особенностей: плотностью (количество особей на единице пространства), адаптивностью (способностью осваивать новые субстраты за счет фонда дремлющих форм, физиологической адаптации активной микрофлоры и др.). Популяции (не только особи!) обладают возрастом, стареют. Микробные ценозы могут быть неполноценными по составу — не содержать некоторых специализированных групп микробов. Важным свойством микробных ассоциаций является их способность к авторегуляции состава и деятельности.
К числу регуляторных механизмов микробных ассоциаций относится характер взаимоотношений между микробными популяциями. Он бывает различным и сложным.
Широко распространены в природе антагонистические взаимоотношения и подавление продуцентами антибиотиков (в первую очередь грибами и актиномицетами) других организмов и изменение кислотности среды в зонах обитания организма при разложении органического вещества.
Антагонизм может быть и более драматическим. Хищники (простейшие) выедают бактерии и водоросли в значительных количествах. Взаимосвязь между хищниками и жертвами оказывается очень прочной: при падении числа особей жертв отмирает часть популяции хищника и, как следствие, возрастает численность клеток жертв, а затем и хищника. Устанавливается своеобразный пульсирующий характер взаимодействия.
Другим примером антагонистических взаимоотношений является паразитизм. В роли паразитов выступают внутриклеточные паразитические бактерии из рода Bdellovibrio и вирусы бактерий — бактериофаги.
Известны многочисленные примеры паразитирования бактерий на мицелии грибов, хищных грибов, активно захватывающих и лизирующих мелких почвенных червей (нематод).
Особым случаем взаимодействия микроорганизмов — крайнее проявление симбиоза — являются лишайники. Они представляют собой ассоциацию из водорослей и грибов. Часто им сопутствуют бактерии. Эти ассоциации очень устойчивы, рассматриваются в специальном разделе, но, по сути дела, являются микробными.
Сложный характер взаимодействия двух и тем более многих компонентов ассоциаций оказывает большое влияние на микробные ценозы, особенно при усилении влияния отдельных групп и последующей смены доминирующих организмов.
Другим источником воздействий на микрофлору почв является сам субстрат. Хорошо известны явления токсичности почв за счет кислой реакции, наличия токсических соединений алюминия и др. Ученые исследуют также явление микробостазиса — свойства почвы задерживать рост и развитие обитающих в ней микробов. Механизмы этих процессов не изучены, но, по-видимому, связаны со специфическим влиянием продуктов жизнедеятельности микроорганизмов и веществ, попадающих в почву при отмирании и лизисе клеток. Почва как своеобразное «живое тело» сопротивляется избыточному развитию микроорганизмов, поддерживая их в дремлющем (латентном) состоянии и «разрешая» вспышки фаворитных групп, обеспеченных в краткие промежутки времени пищей и энергией.
Ризосерная микрофлора
Высшие растения, являясь основным источником питательных веществ для преобладающего числа микробного населения почв — гетеротрофов,— оказывают существенное влияние на микробные ценозы.
Зоны, непосредственно примыкающие к корням живых растений, являются областями активного развития микроорганизмов. Это связано прежде всего с выделениями из корней (экзосмосом) органических веществ, синтезированных растениями. Совокупность микроорганизмов, содержащихся в большом количестве в узкой зоне вокруг корней, называют ризосферной микрофлорой, а саму зону — ризосферой. Кроме того, существует представление о ризоплане — непосредственной поверхности корня, заселенной микробами. Ясно, что метаболизм (обмен веществ) корней оказывает большое влияние на почвенную среду, прилегающую к корням. Считают, например, что корни увеличивают кислотность примыкающих к ним микрослоев почвы за счет выделения углекислоты и Н+ ионов. Такие изменения возможны в пределах нескольких миллиметров вокруг корня. Важным источником стимуляции почвенного микронаселения является выделение корнями питательных веществ. Патогенные и симбиотические микроорганизмы привязаны к ним либо способны растворять стенку клеток корня и проникать внутрь цитоплазмы. Экзосмос органических веществ из корней растений обусловлен активными процессами, пассивной диффузией или выделениями из отмирающих клеток.
Молодые корешки обычно покрыты слизистыми чехликами, обильно заселенными микробами.
В продуктах экзосмоса корней обнаружено большое количество различных веществ, в том числе 10 разных Сахаров, 23 аминокислоты, 10 витаминов, полисахаридные слизи, органические кислоты и др. Характер выделений зависит от вида и возраста растений. К сожалению, еще нет достаточных сведений о процессах корневого экзосмоса и использования веществ микроорганизмами в условиях природной нестерильной среды.
Сфера воздействия корней на микрофлору в почве определяется лишь приблизительно но увеличению числа микробов по мере приближения к поверхности корня. Большинство трупп микроорганизмов обнаруживается в большем числе в ризосфере (Р), чем в окружающей почве (П), что можно выразить отношением: Р/П.
Иллюстрации к этому положению приведены в следующей таблице:
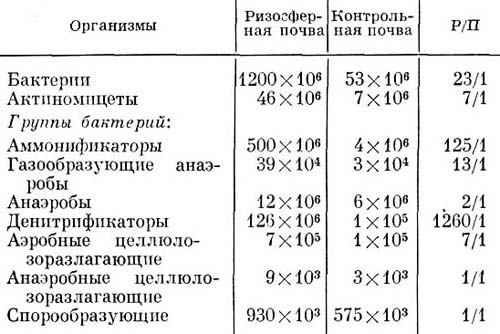
Сравнение числа разных бактерий и актиномицетов в ризосфере пшеницы и в контрольной почве
Не все бактерии одинаково реагируют на стимулирующее действие корней. Так, грамотрицательные бактерии явно лучше развиваются в ризосфере, чем грамположительные неспорообразующие бактерии. Способность бактерий заселять зону корня связана не только с выделяемым веществом, но и с отношением к физическим факторам, антагонизму. Многие исследователи обнаружили, что бактерии, способные разлагать целлюлозу, растворять фосфаты, использовать белки и сахара, синтезировать витамины и бактериальные полисахариды,— являются типичными обитателями ризосферы. Удается также отметить некоторую корреляцию между таксономическими и физиологическими группами бактерий в ризосфере. Так, например, виды Pseudomonas доминируют в ризосфере, а виды Arthrobacter — в окружающей почве. Представители обоих родов заметно различаются по потребностям в ростовых факторах.
Меньше внимания уделялось актиномицетам, хотя их приуроченность к ризосфере установлена. Возможно, что в этих условиях они образуют активные вещества (антибиотики), угнетающие развитие патогенов на корнях. В этом вопросе остается много неясностей. По видовому составу и особенно физиологии виды актиномицетов, обитающие в ризосфере и в почве, не отличаются.
На ризосферную микрофлору влияют вид, возраст растений и их состояние, положение и характер распределения корней, тип почвы и окружение. Корни растений стимулируют или угнетают микробов в разной степени. Бобовые растения чаще всего стимулируют развитие микробов. В ризосфере клеверов, например, обнаруживается значительно больше микроорганизмов, чем в зоне корней злаков и деревьев. Корневые выделения растений в случае длительного выращивания одних и тех же культур растений (монокультур) на одних и тех же площадях приводят к так называемому «почвенному утомлению». Такая обстановка в сочетании с монотонным по составу растительным опадом вызывает селекцию отдельных групп и даже видов микроорганизмов и их чрезмерное развитие в почвах. Следствием этого являются стойкие заболевания растений (при развитии патогенных для растений микробов), уносящие урожай. В пределах поверхности одного корешка микрофлора оказывается разной и обилие микроорганизмов нарастает к кончикам корней, где, например, отмечен наибольший экзосмос аминокислот.
Ризосферный эффект увеличивается после прорастания семени и достигает максимума в период цветения и плодоношения растений. Таким образом, возраст и старение растений играет большую роль в формировании и деятельности ризосферной микрофлоры. Представление об этом дано в таблице:
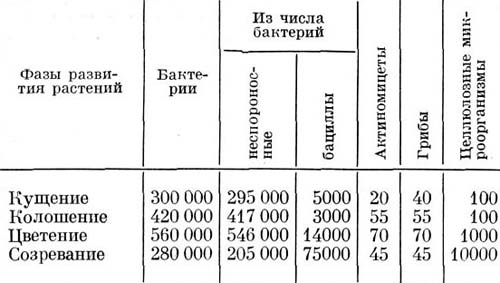
Групповой состав и численность микрофлоры ризосферы пшеницы (чило микроорганизмов в тыс. на 1 г почвы). По Е.Н. Мишустину, 1972 г.
Степень освещенности и температура воздуха также влияют на корневые выделения и, следовательно, на микроорганизмы.
Ризосферный эффект более ярко выражен в песчаных почвах и менее — в гумусных. В пустынных районах ризосфера является, по-видимому, единственной зоной, где активно развивается микрофлора. В любой почве изменения окружающей среды, включая агротехнические мероприятия, оказывают меньшее воздействие на микроорганизмы в ризосфере по сравнению с обитателями почвы. Ризосферная зона представляет собой своеобразную «буферную» систему, препятствующую воздействию среды на микрофлору.
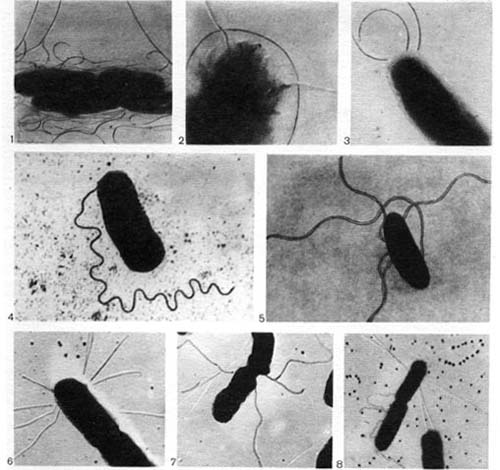
Таблица 42. Жгутики (1—5) и жгутикоподобные образования (6-—8) у клеток клубеньковых бактерий: 1, 3, 6—8 — из клубеньков клевера (увел, х 15 000—25 000); 2 — люцерны (увел, х 40 000); 4 — фасоли (увел, около 25 000, по Дж. Зильгеру); 5 — гороха (увел, около 25 000, по Дж. Лею, А. Расселу).
Мероприятия, проводимые человеком, оказывают огромное воздействие на микробные ассоциации. Среди них наиболее существенно применение химических средств борьбы с сорняками (гербициды), всевозможных протравителей семян, минеральных удобрений. Все это в сочетании с разными типами обработки почв (вспашка, орошение, мелиорация) изменяет микробные ценозы, часто стойко и не всегда в благожелательном для хозяйства направлении. В этом плане во всем мире ведется большая исследовательская работа.
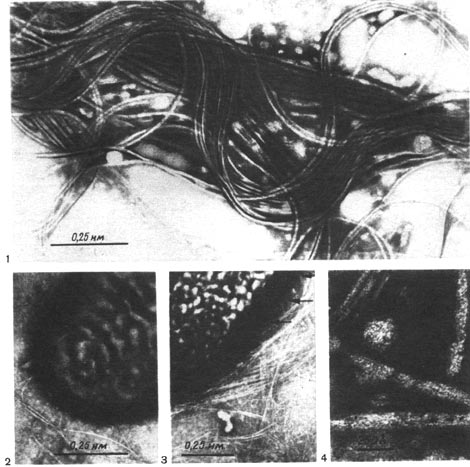
Таблица 43. Жгутики (1) и нитевидные выросты — фимбрии (2—4) у клеток клубеньковых бактерий люпина. Стрелками показаны места выхода фимбрии из клетки (по Майеру).
Широко распространенное, даже среди некоторой части специалистов, мнение о почве как о подобии питательной среды, которую можно заселить любыми микроорганизмами, глубоко неверно. Колонизация микробами отдельных участков (как было показано, часто очень мелких) может иметь место в течение непродолжительного времени. Затем вступают в действие те или иные регуляторные механизмы, и популяции «пришельцев» оказываются сведенными к минимуму. Настойчивые попытки на протяжении многих лет применить бактеризацию (внесение) почвы микроорганизмами с полезными для растений свойствами (фиксаторов азота, активных минерализаторов фосфорсодержащих веществ) оказались неуспешными.
Колонизация почвы микроорганизмами (в том числе не содержащимися в ассоциациях в данном участке) все же ограниченно возможна, если не лимитируется следующими условиями:
а) наличием пригодного места в данный момент (в реальных микрозонах обитания микроорганизмов, а не вообще в почве);
б) присутствием в достаточном количестве субстрата, обеспечивающего длительное развитие данного организма;
в) способностью использовать многие источники энергии и пищи в экосистеме, и причем более эффективно, чем аборигенная микрофлора;
г) высокой толерантностью (выносливостью) попадающего организма к микроэкологическим факторам — колебаниям рН и температуры, осмотическому давлению, содержанию кислорода, окислительно-восстановительному потенциалу, влажности и другим, обеспечивающим вегетативный рост;
д) наличием механизмов активного воздействия на другие организмы — образованием физиологически активных метаболитов, способностью к антагонизму и устойчивостью к продуктам жизнедеятельности других организмов.
Несоблюдение указанных условий и даже некоторых из них делает невозможным активное поселение микроорганизмов в почве. Более того, как уже отмечалось, входящие в состав ценоза микробы при отсутствии возможностей приспособиться к изменившимся условиям среды выпадают из активного участия в деятельности ценоза, переходя в переживающее состояние.
Каждый вид микроорганизма способен расти, развиваться и размножаться в рамках внешних условий, которые отражают их уровень толерантности или экологическую амплитуду. Эти рамки определены критическими величинами факторов. Отдельные организмы способны существовать при крайних (экстремальных) значениях факторов среды и часто становятся узкоспециализированными — облигатными (обязательными) по отношению к уровню действующего фактора. Таковыми являются облигатные галофилы (род Halobacterium), растущие в насыщенных растворах солей, многие облигатные термофилы, глубоководные барофильные бактерии (устойчивые к высокому давлению), выдерживающие давление 1400 атм, облигатные анаэробы, погибающие при незначительных примесях кислорода в атмосфере (род Selenomonas и др.). Ряд бактерий и грибов способны размножаться при О °С, если водный раствор не замерзает (растворы солей, в отличие от чистой воды, замерзают при более низкой температуре).
Имеются многочисленные примеры исключительности микробного мира в целом по отношению к факторам среды в экстремальном (крайнем) выражении. Так, например, опасным температурным пределом для животных, включая простейших, является 50 °С, максимум для грибов 56—60 °С. Сине-зеленые водоросли (род Synechococcus) активны в горячих источниках при 73—75 °С, а некоторые флексибактерии активно размножаются в горячих гейзерах (90 °С). Зона толерантности микробного мира поистине грандиозна, ее границы часто находятся на предельных значениях отдельных факторов. Эта особенность микроорганизмов обеспечивает им практически беспредельное развитие на нашей планете.
Большинство же микроорганизмов ограничено более узкими рамками, и их развитие либо задерживается, либо в результате воздействия отдельных факторов наступает гибель и разрушение клеток. Большую роль в выживаемости микроорганизмов в условиях, которые могут оказаться губительными, играют «факторы компенсации». Так, например, температурный барьер преодолевается микробами, не приспособленными к этому, при наличии питательных элементов. Термофильная бактерия Bacillus stearothermophilus развивается при низкой температуре при наличии в среде ростовых факторов. Arthrobacter globiformis толерантен к высокой и низкой температуре в присутствии солей (NaCl). Низкая температура придает Bacillus stearothermophilus нечувствительность к токсинам, не мешая ее росту.
Нет практически ни одного вещества (в первую очередь содержащего углерод), которое не могло бы быть разложено микроорганизмами. Самые устойчивые соединения — асфальты, битумы и новые синтезированные химическим путем соединения, не встречающиеся в природе, также атакуются микробами.
Все изложенные выше сведения, составляющие лишь фрагменты того, что известно современной микробиологии, дают основание самому читателю сделать вывод об огромной и исключительной роли микроорганизмов в круговороте веществ в природе.



