




Жизнь растений
Бактерии и актиномицеты
Общая характеристика бактерий и актиномицетов
Бактерии (от слова bacterion — палочка) — это наиболее широко распространенная в природе группа микроорганизмов, представляющих собой большой и чрезвычайно разнообразный мир микроскопических существ. Клетки наиболее мелких шаровидных бактерий имеют в поперечнике менее 0,1 мкм (т. е. 0,0001 мм). Подавляющее большинство бактерий — это палочки, толщина которых в среднем составляет 0,5—1 мкм, а длина 2—3 мкм. Очень редко встречаются бактерии-«гиганты», клетки которых имеют в диаметре 5—10 мкм, а в длину достигают 30—100 мкм.
Крайне малые размеры клеток являются характерной, но не главной особенностью бактерий. Все бактерии представлены особым типом клеток, лишенных истинного ядра, окруженного ядерной мембраной. Аналогом ядра у бактерий является нуклеоид — ДНК-содержащая плазма, не отграниченная от цитоплазмы мембраной. Кроме того, для бактериальных клеток характерны отсутствие митохондрий, хлоропла-стов, а также особое строение и состав мембранных структур и клеточных стенок. Организмы, в клетках которых отсутствует истинное ядро, называются прокариотами (доядер-ными) или протоцитами (т, е. организмами с примитивной организацией клеток).
Бактерии, в широком смысле слова,— это прокариотные организмы. К прокариотам относятся такие группы микроорганизмов, как эубактерии, спирохеты, микоплаз-мы, миксобактерии, лучистые грибки (актино-мицеты) и сине-зеленые водоросли (цианобактерии). Форма клеток у бактерий может быть не только палочковидной (цилиндрической), но и шаровидной (кокки), спиральной (вибрионы, спириллы, спирохеты). Актиномицеты же и родственные им организмы образуют длинные ветвящиеся клетки — гифы, формирующие мицелий (сплетение гиф). Клетки микоплазм, лишенные плотной оболочки, способны принимать самые причудливые, постоянно изменяющиеся формы.
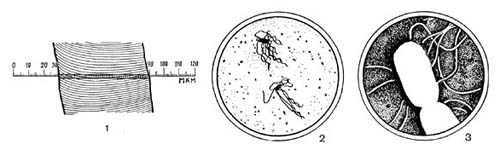
Рис. 1. Сравнительная величина волоса и бактерий: 1 — увеличенное изображение волоса (X 500); 2 — бактерии в поле зрения светового микроскопа (X 2000); з — бактерии под электронным микроскопом (X 20 000).
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель XVII в., впервые создавший совершенную лупу-микроскоп, увеличивавший предметы в 160 — 270 раз. Со времен Левенгу-ка техника исследования микробиологических объектов шагнула далеко вперед. Созданы световые микроскопы, увеличивающие объекты в 2000 и более раз. С помощью современного электронного микроскопа, увеличивающего предметы в 200 000 — 500 000 раз, можно различать и изучать самые мелкие микроорганизмы. Для сравнения на рисунке 1 показаны размеры волоса в микрометрах, изображение бактерии, полученное в световом микроскопе, и часть той же бактерии в поле зрения электронного микроскопа.
Вооруженная усовершенствованными приборами, микробиология все глубже и полнее познает свойства и особенности мира невидимых. Ученые-микробиологи разрабатывают новые методы наилучшего использования полезных микробов и пресечения деятельности вредных.
Разработанный русским микробиологом С. Н. Виноградским метод выращивания микробов в лабораториях на элективных (избирательных) питательных средах дал возможность более подробно изучить различные микроорганизмы и их распространение в природе. Используя этот метод, ученые установили, что бактерии обнаруживаются всюду: в каждой капле даже самой чистой воды, в крупинках почвы, в воздухе, на скалах Земли Франца-Иосифа и в снегах полярных областей, в океане на Северном полюсе. Разнообразные виды бактерий найдены также в почвах пустыни Сахара, в грунте, взятом со дна океана на глубине 4 км, и в нефти, добытой из глубоко залегающих нефтяных пластов. Бактерии способны жить даже в воде горячих источников с температурой около 80° С.
Благодаря ничтожным размерам бактерии легко проникают в трещины, щели, поры. Они очень выносливы и приспособлены к неблагоприятным условиям существования: переносят высушивание, сильные холода, нагревание до 80 — 90° С, не теряя при этом жизнеспособности. А споры бактерий выдерживают даже кипячение.
Бактерии играют важную роль в народном хозяйстве и в быту человека. Велика их роль в почвообразовательных процессах. Бактерии нашли широкое применение в растениеводстве и животноводстве, в различных отраслях пищевой промышленности. Микробиология все шире внедряется в текстильную и кожевенную отрасли промышленности. Такие производства, как мочка льна, обработка кожи, не могут обойтись без использования продуктов жизнедеятельности бактерий.
Необычайно велико значение бактерий в патогенезе заболеваний человека, животных и растений.
Почвенные бактерии оказывают большое влияние на рост и развитие растений. Вокруг корней и на корнях растений развивается большое количество различных видов бактерий, причем не всех видов, а только определенных, специфичных для каждого вида растений. Эта микрофлора может быть полезной или вредной для растения в зависимости от окружающей среды и состояния самого растения.
В процессе жизнедеятельности бактерии образуют ряд соединений, которые широко используются человеком: антибиотики, аминокислоты, витамины, ферменты и другие соединения. Много ценнейших биологически активных веществ, продуцируемых микроорганизмами, нам пока неизвестно. С каждым годом список этих соединений пополняется; мир микробов — неисчерпаемый источник различных важных веществ.
Громадное значение имеет геологическая деятельность бактерий. Бактерии принимают самое активное участие в круговороте веществ в природе. Все органические соединения и значительная часть неорганических подвергаются при этом существенным изменениям. И этот круговорот веществ является основой существования жизни на Земле.
Настоящий том посвящен прокариотным организмам (за исключением сине-зеленых водорослей, которые будут описаны в 3-м томе). Хотя раньше бактерии относили к растительным организмам, в настоящее время показано, что бактерии как прокариоты составляют особое царство живых существ, отличное от царств растений и животных. Следуя традиции, редакция начинает издание «Жизнь растений» с описания бактерий как низших организмов — протоцитов. В первой части раздела по бактериям (введении) кратко излагаются основные принципы классификации, строение клеток и обмен веществ у бактерий. Во второй части приводятся сведения об основных систематических группах бактерий. Заключают раздел главы, описывающие экологию, физиологические группы и биохимические особенности бактерий.
Основные принципы классиффикации бактерий и актиномицетов
Классификация живых существ является одним из наиболее трудных разделов биологической науки. В ней, как в фокусе, концентрируются все наши познания об организмах. Чем глубже и полнее наши сведения об организмах, тем точнее мы их классифицируем. С прогрессом биологической науки совершенствуется и классификация живых существ.
Систематика низших организмов совершенствуется крайне слабо. Объясняется это значительной бедностью морфологических и цитологических признаков у микробов, а также трудностями в изучении филогенеза этих существ.
В классификации бактерий существуют два направления. Первое — каталогизация форм — производится на основе какого-либо одного или нескольких признаков, часто случайных, не связанных между собой. Второе — это построение системы на основе филогенетических данных, полученных путем комплексного, всестороннего изучения организмов.
По мере изучения биологии бактерий исследователи начали применять для классификации, помимо морфологических, многие другие признаки: физио лого-биохимические, цитологические, серологические, иммунологические и др. В современных классификациях авторы используют любой признак, лишь бы он выделялся и давал возможность распознать изучаемый организм.
Специализация биохимической деятельности микроорганизмов побуждает некоторых исследователей подразделять бактерии на отдельные физиологические группы. Наиболее ярко выражена такая группировка в классификациях Орла-Иенсена и Берги. Бактерии подразделяются на основе их способности вызывать заболевания у человека, животных и растений; формируются особые группы патогенных и фитопатогенных форм только по одному этому признаку.
Не удивительно, что с каждым новым изданием определителя микробов меняется группировка микроорганизмов. Микроорганизмы перемещаются из одной таксономической группы в другую, причем перемещаются не только отдельные виды, но и роды и даже более высокие систематические единицы. В одной и той же группе организмы распределяются то по морфологическим признакам, то по физиологическим; к одной и той же группе относятся подвижные и неподвижные формы, грамположительные и грамотрицательные бактерии.
Невольно возникает вопрос: возможна ли классификация микробов не на формалистических принципах, не в целях каталогизации и диагностики, а на принципе филогенетических взаимоотношений, на основе родства изучаемых организмов? Можно ли строить филогенез бактерий подобно тому, как это осуществляется в классификации растительного мира?
Известно, что за последние десятилетия в области филогенетической систематики растений достигнуты большие успехи. Систематика растений строится на основе комплексных исследований различных дисциплин биологической науки: морфологии, цитологии, генетики, биохимии, серологии, геоботаники, палеоботаники и др.
Особенности строения бактерий не позволяют при их изучении применять все те методы, которые с успехом используются в ботанике. Простота строения бактерий, отсутствие типичного ядра и других органоидов в их клетках, а также полового процесса исключают .возможность использования цитоморфологического метода в той степени, как это имеет место в систематике растений. Исключается здесь и исторический метод. Нам почти ничего не известно о далеком прошлом бактерий. Имеющиеся указания на окаменевшие остатки бактерий малочисленны и немного дают для разъяснения исторических процессов эволюционного развития этих организмов.
Бактерии широко распространены в природе, характерной их особенностью является космополитизм. Имеющиеся данные о географическом распределении и специфике бактерий пока не дают возможности использовать их в целях систематики.
Возможный и желательный биохимический метод систематики, к сожалению, применительно к микробиологии остается пока слабо разработанным. Однако некоторые биохимические работы представляют значительный интерес, так как дают основания для подразделения бактерий на группы и виды.
Современные классификации и схемы филогенеза организмов, несмотря на их умозрительность, побуждают исследователей по-новому относиться к накопленному экспериментальному материалу и систематизировать его с определенных позиций, а не по случайным признакам.
Быстрое размножение и развитие бактерий на питательных средах дают возможность проводить широкие эксперименты и получать результаты в короткий срок. Особенно это важно при массовом анализе большого числа поколений микроорганизмов. Как бы ни были скудны морфологические признаки у бактерий и лучистых грибков, все же в своей совокупности и при сравнительном анализе они с успехом используются в микробиологии для установления родства микроорганизмов.
Следует подчеркнуть, что один и тот же внешний морфологический признак микробов может иметь разное значение. Например, существенный для микобактерий признак ветвления клеток вовсе не характерен для таких истинных бактерий, как уксуснокислые бактерии, азотобактер и др. У микобактерий ветвление — нормальный формообразовательный процесс; у бактерий ветвление — патологический процесс, наблюдаемый при дегенеративном перерождении, ветвящиеся клетки у этих бактерий представляют инволюционные, отмирающие формы.
Наиболее приемлемый метод для выявления родства микробов — метод экспериментальной изменчивости. С помощью этого метода удалось выявить родство у некоторых бактерий и микобактерий, установить близость микококков с микобактериями, микобактерий с проактиномицетами. а проактиномицетов с актиномицетами.
Культуры актиномицетов и бактерий при длительном нахождении в субстратах, в которых имеется какой-либо искусственный источник питания, например сахар мальтоза, образуют штаммы, сбраживающие мальтозу. У сбраживающего мальтозу вида микроба появляется фермент, расщепляющий мальтозу. При истощении субстрата этот фермент утрачивается.
Такие изменения описаны у очень многих микробов: бактерий, актиномицетов, дрожжей и грибов. Имеются данные, показывающие, что изменения подобного типа связаны с генетическим аппаратом клетки.
Разрешение вопроса, связанного с определением вида, важно не только с позиции чисто познавательной, общебиологической, но и с рядом прикладных задач. Особенно велико практическое значение данного вопроса в препаративной микробиологии, инфекционном патогенезе человека, животных и растений, в технической и бродильной микробиологии и других областях. В промышленности и сельском хозяйстве, где действующим фактором являются бактерии, проблема вида и способы определения его имеют большое значение. Малейшие неточности в определении вида могут привести к нежелательным последствиям.
Решение важнейших биологических вопросов, таких, как изменчивость и наследственность, также тесно связано с определением вида. Практическая значимость данной проблемы побуждает специалистов все более углубленно заниматься ее разрешением. В настоящее время проблемой вида занимаются многочисленные лаборатории во всех странах мира.
В классификации организмов проблема вида является самой важной и трудной. Без определения вида нельзя строить классификацию. В бактериологии вид как основная систематическая единица остается весьма неопределенным. Трактовка вида разноречива, часто в зависимости от личных взглядов и вкусов специалиста в нее вносится много субъективного, специфического.
При изучении мира бактерий исходным положением должно быть признание таксономических единиц, прежде всего признание наличия видов как объективной реальности, существующей в природе.
Вид представляет собой продукт эволюции живой материи, имеющий свою историю развития, формирования и стабилизации в результате приспособления к условиям существования.
Методы познания вида могут меняться и уточняться по мере изучения биологии организма и развития науки о жизни. От линнеев-ского понимания вида как стабильной единицы до понимания вида Дарвином и современными биологами как развивающегося естествознание прошло большой путь.
Надо отметить, что для установления вида нельзя ограничиваться каким-либо одним признаком. Только по совокупности признаков -морфологических, культуральных, цитохимических, физиолого-биохимических и др.— можно охарактеризовать вид. Вид у бактерий определяется суммой разнообразных признаков и свойств.
Установлению вида у бактерий существенно помогает то, что этих микробов можно культивировать на искусственных питательных средах в чистом, изолированном виде и при этом наблюдать за развитием не только отдельных клеток, но и всей популяции в целом, отмечая любые видимые изменения на всех стадиях развития. Быстрый рост бактерий дает возможность разрешать многие вопросы, связанные с изменчивостью организма, его наследственной стабильностью, и вместе с этим устанавливать ведущие видовые признаки. Сопоставление культур в процессе их развития на определенных средах дает возможность выявить сходство или различие между ними.
При группировке бактерий мы придаем большое значение физиологическим показателям как систематическим признакам. Например, азотфиксаторы определяются по азотфиксации, клетчаткоразрушающие бактерии — по способности разлагать клетчатку и т. д. Большую ценность имеют биохимические показатели: образование антибиотиков, спиртов, уксусной кислоты, молочной кислоты и др.
Некоторые виды микробов образуют различные пигменты: красные, синие, зеленые, оранжевые, бурые, черные и смешанных цветов. Большинство микробов не образуют пигментов, их колонии бесцветны. Пигменты — признак стабильный, который присущ определенным видам микробов. Этот признак может быть использован и в систематике бактерий. Биологическое значение пигментов пока малоизвестно.
Виды микроорганизмов подразделяются по совокупности морфологических, физиологических и биохимических признаков. У каждого организма много признаков, но не каждый признак может быть использован в систематике. Имеются признаки ведущие, характеризующие вид, и признаки соподчиненные. Ведущим признаком могут быть различные показатели у разных групп бактерий. Для одних видов таковыми является способность ассимилировать углеводы, для других — образовывать антибиотики, для третьих — сбраживать сахар или синтезировать специфические метаболиты, пигменты, кислоты, гормоны и т. д. У некоторых видов ведущим признаком является способность образовывать клубеньки на корнях или на листьях растений. Некоторые виды подразделяются по своим ферментативным свойствам, например уксуснокислые, молочнокислые, пропионовокислые и другие бактерии. Многие виды подразделяются по вирулентности и патогенности в отношении растений, животных и человека.
В последние годы большое внимание уделяется химическому составу клеток как дифференцирующему признаку. Одни исследователи предлагают подразделять виды по химическому составу клеточных оболочек, другие — по составу белков протоплазмы или нуклеиновых кислот, третьи — по составу только дезок-сирибонуклеиновой кислоты.
Следует отметить, что в последнее время изучается рекомбинация отдельных видов бактерий и актиномицетов. Это явление напоминает гибридизацию у высших организмов. Установлено, что при контакте клеток (чаще дефектных) двух разных штаммов бактерий или актиномицетов свойства одного штамма переходят к другому. В результате получаются смешанные формы с признаками двух исходных культур. Такой процесс происходит между двумя родственными организмами.
В микробиологии может с успехом применяться филогенетический метод определения вида. Метод экспериментальной изменчивости дает возможность установить не только вариабильность и полиморфизм культур, но и по образовавшимся вариантам судить о родстве сопоставляемых культур.
В медицинской бактериологической практике широко применяется серологический метод (метод, с помощью которого изучаются антитела и связанные с ними реакции иммунитета - обезвреживание токсинов, агглютинация микробов и др.) для диагностицирования патогенности бактерий. Этот метод очень чувствителен, с его помощью выявляются штаммы, очень близкие между собой.
Тем не менее серологический метод не используется для систематики бактерий в той мере, как это следовало бы ожидать. В большинстве случаев при помощи его не удается выявлять или разделять виды. Многочисленные попытки в этом направлении дают противоречивые результаты, явно не согласующиеся с показателями, полученными другими методами. Серологический метод наиболее хорошо разработан для дифференцирования бактерий кишечной группы.
Для идентификации видов бактерий и актиномицетов микробиологи пытаются применить метод фаголизиса. Он основан на способности фагов лизировать (разрушать) клетки определенных видов бактерий. Имеются поли- и моновалентные фаги. Для дифференцирования видов пригодны только моновалентные фаги, которые обладают строгой специфичностью.
В фитопатологии для установления видов применяют паразитологический и токсикологический методы. Известно, что многие паразитические организмы — бактерии и грибы -строго специализированы в отношении выбора хозяина, поражают только определенные виды растений. У некоторых микроорганизмов эта специфика настолько тонкая и постоянная, что по ней можно определить вид.
Важным видовым показателем, по нашим наблюдениям, является специфика антагонизма. Микробы-антагонисты обладают способностью подавлять рост других микробов продуктами метаболизма - - антибиотическими веществами. Антагонизм между культурами одного и того же вида не наблюдается. Этот признак весьма показателен, строго постоянен и легко воспроизводим. При помощи метода, в котором используется антагонизм микроорганизмов, удается выявить виды, не дифференцирующиеся другими способами.
В ботанической литературе часто употребляются термины «поливалентные» и «моновалентные» виды. Моновалентные виды представлены единичными формами или одной-единственной культурой, а поливалентные состоят из большого числа соподчиненных единиц: разновидностей, вариантов, форм и формирований.
У многих видов бактерий поливалентность видов не обнаружена вследствие того, что мы не имеем достаточно основательных признаков для установления видов и, следовательно, разновидностей, форм и вариантов. Возможно, что у бактерий вообще нет моновалентных видов, состоящих из одного штамма. Более широко распространены в природе поливалентные виды.
Многие поливалентные формы по количественному составу и разнообразию форм, составляющих их, выходят за рамки объема вида и становятся надвидами, т. е. такими таксономическими группами, которые занимают промежуточное положение между родом и видом.
Итак, в мире бактерий существуют совершенно определенные виды как реально существующие таксономические единицы, а не вымышленные схемы. Понятие вида мы можем сформулировать так: вид — это группа близких между собой организмов, имеющих общий корень происхождения и на данном этапе эволюции характеризующихся определенными морфологическими, биохимическими и физиологическими признаками, обособленных отбором от других видов и приспособленных к определенной среде обитания.
Строение и химический состав бактерильной клетки
Общая схема строения бактериальной клетки показана на рисунке 2. Внутренняя организация бактериальной клетки сложна. Каждая систематическая группа микроорганизмов имеет свои специфические особенности строения.
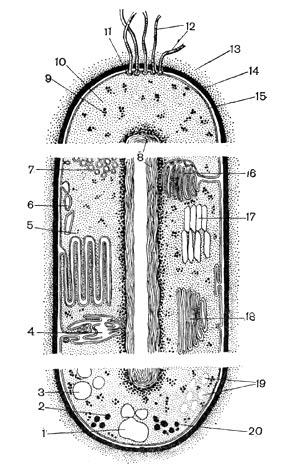
Рис. 2. Схематическое изображение строения бактериальной клетки (по Г. Шлегелю): I — гранулы поли-В-оксимасляной кислоты; 2 — жировые капельки; з — включения серы; 4 — трубчатые тилакоиды; S — пластинчатые тилакоиды; в — пузырьки; 7 — хроматофоры; g — ядро (нуклеоид); 9 — рибосомы; 10 — цитоплазма; и — базальное тельце; 12 — жгутики; 13 — капсула; 14 — клеточная стенка; 15 — цитоплазматическая мембрана; 16 — мезосома; П — газовые вакуоли; is — ламеллярные структуры; 19 — гранулы полисахарида; 20 — гранулы полифосфата. Основные структуры бактериальной клетки представлены в верхней части рисунка, дополнительные, мембранные структуры, имеющиеся у фототрофных и нефототрофных бактерий,— в средней части, а включения запасных веществ — в нижней части.
Клеточная стенка
Клетка бактерий одета плотной оболочкой. Этот поверхностный слой, расположенный снаружи от цитоплазматиче-ской мембраны, называют клеточной стенкой (рис. 2, 14). Стенка выполняет защитную и опорную функции, а также придает клетке постоянную, характерную для нее форму (например, форму палочки или кокка) и представляет собой наружный скелет клетки. Эта плотная оболочка роднит бактерии с растительными клетками, что отличает их от животных клеток, имеющих мягкие оболочки. Внутри бактериальной клетки осмотическое давление в несколько раз, а иногда и в десятки раз выше, чем во внешней среде. Поэтому клетка быстро разорвалась бы, если бы она не была защищена такой плотной, жесткой структурой, как клеточная стенка.
Толщина клеточной стенки 0,01—0,04 мкм. Она составляет от 10 до 50% сухой массы бактерий. Количество материала, из которого построена клеточная стенка, изменяется в течение роста бактерий и обычно увеличивается с возрастом.
Основным структурным компонентом стенок, основой их жесткой структуры почти у всех исследованных до настоящего времени бактерий является муреин (гликопептид, муко-пептид). Это органическое соединение сложного строения, в состав которого входят сахара, несущие азот, — аминосахара и 4—5 аминокислот. Причем аминокислоты клеточных стенок имеют необычную форму (D-стереоизомеры), которая в природе редко встречается.
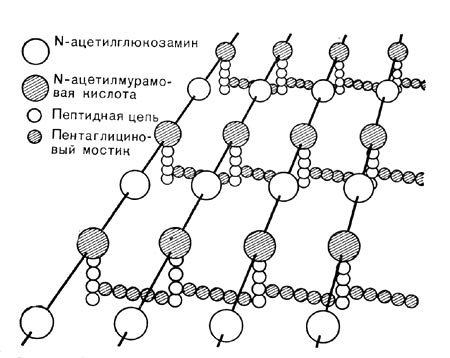
Рис. 3. Схематическое изображение структуры глико-пептида клеточной стенки. К полисахаридным цепочкам, состоящим из JV-ацетилглюкоз-амина и IV-ацетилмурамовой кислоты, присоединены цепочки из аминокислотных единиц — пептиды. Пептиды, связанные друг с другом пеитаглициновыми мостиками, образуют поперечные связи полисахаридных цепочек.
Составные части клеточной стенки, ее компоненты, образуют сложную прочную структуру (рис. 3, 4 и 5).
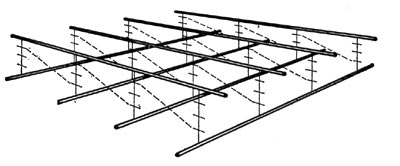
Рис. 4. Расположение макромолекул гликопептида клеточной стенки. Жирными линиями показаны скелеты полисахаридных цепей, расположенные ближе к поверхности; более тонкими — подстилающие их полисахаридные цепи. Линии с поперечными штрихами изображают пептидные цепочки, связывающие эти полисахаридные цепи. Межпептидные мостики, состоящие из пяти остатков глицина, обозначены пунктирными линиями.
С помощью способа окраски, впервые предложенного в 1884 г. Кристианом Грамом, бактерии могут быть разделены на две группы: грамположительные и грамотрицательные. Грамположительные организмы способны связывать некоторые анилиновые красители, такие, как кристаллический фиолетовый, и после обработки иодом, а затем спиртом (или ацетоном) сохранять комплекс иод-краситель. Те же бактерии, у которых под влиянием этилового спирта этот комплекс разрушается (клетки обесцвечиваются), относятся к грамотрицательным.
Химический состав клеточных стенок грам-положительных и грамотрицательных бактерий различен.
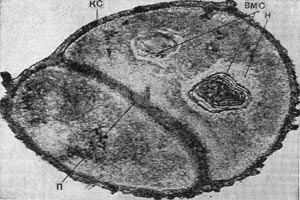
Рис. 5. Тонкое строение клеток грамположптельных кокков — Micrococcus aurantiacus. кс — клеточная стенка; п — перегородка; вмс — внутри-цитоплазматические мембранные структуры; н — нуклеоид. Увел. х65 000.
У грамположительных бактерий в состав клеточных стенок входят, кроме мукопептидов, полисахариды (сложные, высокомолекулярные сахара), тейхоевые кислоты (сложные по составу и структуре соединения, состоящие из Сахаров, спиртов, аминокислот и фосфорной кислоты). Полисахариды и тейхоевые кислоты связаны с каркасом стенок — мурей-ном. Какую структуру образуют эти составные части клеточной стенки грамположительных бактерий, мы пока еще не знаем. С помощью электронных фотографий тонких срезов (слоистости) в стенках грамположительных бактерий не обнаружено. Вероятно, все эти вещества очень плотно связаны между собой.
Стенки грамотрицательных бактерий более сложные по химическому составу, в них содержится значительное количество липидов (жиров), связанных с белками и сахарами в сложные комплексы — липопротеиды и липополисахариды. Муреина в клеточных стенках грамотрицательных бактерий в целом меньше, чем у грамположительных бактерий. Структура стенки грамотрицательных бактерий также более сложная. С помощью электронного микроскопа было установлено, что стенки этих бактерий многослойные (рис. 6).
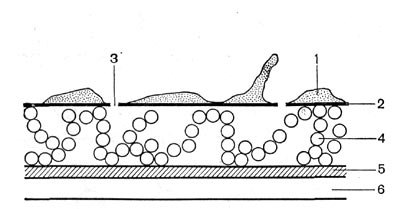
Рис. 6. Схематическое изображение клеточной стенки грамотрицательных бактерий — Bacterium coli (no Роузу): 1 — липопротеидный слой с выступами и бугорками; 2 — липо-полисахаридный слой; з — каналы; 4 — рыхлоупакованные молекулы белка; 5 — гликопептидный слой; в — цитоплазмати-ческая мембрана.
Внутренний слой состоит из муреина. Над ним находится более широкий слой из неплотно упакованных молекул белка. Этот слой в свою очередь покрыт слоем липополисахари-да. Самый верхний слой состоит из липопро-теидов.
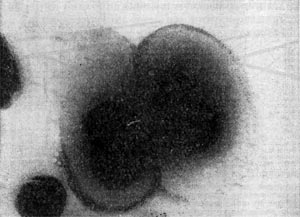
Рис. 7. Капсула клеток Azotobacter chroococcum. Увел. X 25 000.
Клеточная стенка проницаема: через нее питательные вещества свободно проходят в клетку, а продукты обмена выходят в окружающую среду. Крупные молекулы с большим молекулярным весом не проходят через оболочку.
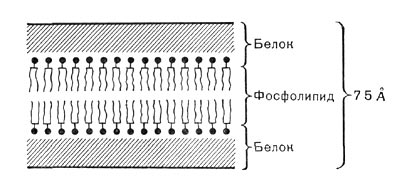
Рис. 8. Схема строения цитоплазматической мембраны.
Капсула
Клеточная стенка многих бактерий сверху окружена слоем слизистого материала — капсулой (рис. 7). Толщина капсулы может во много раз превосходить диаметр самой клетки, а иногда она настолько тонкая, что ее можно увидеть лишь через электронный микроскоп, — микрокапсула.
Капсула не является обязательной частью клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она служит защитным покровом клетки и участвует в водном обмене, предохраняя клетку от высыхания.
По химическому составу капсулы чаще всего представляют собой полисахариды. Иногда они состоят изгликопротеидов (сложные комплексы Сахаров и белков) и полипептидов (род Bacillus), в редких случаях — из клетчатки (род Acetobacter).
Слизистые вещества, выделяемые в субстрат некоторыми бактериями, обусловливают, например, слизисто-тягучую консистенцию испорченного молока и пива.
Цитоплазма.
Все содержимое клетки, за исключением ядра и клеточной стенки, называется цитоплазмой. В жидкой, бесструктурной фазе цитоплазмы (м а т р и к с е) находятся рибосомы, мембранные системы, митохондрии, пластиды и другие структуры, а также запасные питательные вещества. Цитоплазма обладает чрезвычайно сложной, тонкой структурой (слоистая, гранулярная). С помощью электронного микроскопа раскрыты многие интересные детали строения клетки.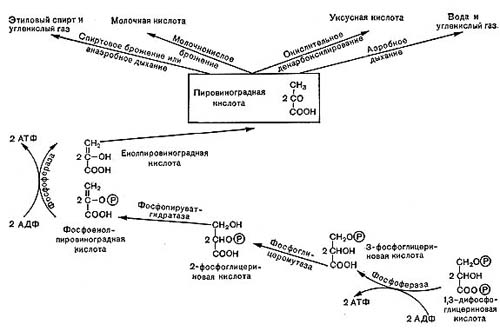
Рис. 9. Основные типы расположения жгутиков и направления движений бактерий.
Внешний липопротеидный слой протопласта бактерий, обладающий особыми физическими и химическими свойствами, называется цитоплазмат и ческой мембраной (рис. 2, 15).
Внутри цитоплазмы находятся все жизненно важные структуры и органеллы.
Цитоплазматическая мембрана выполняет очень важную роль — регулирует поступление веществ в клетку и выделение наружу продуктов обмена.
Через мембрану питательные вещества могут поступать в клетку в результате активного биохимического процесса с участием ферментов. Кроме того, в мембране происходит синтез некоторых составных частей клетки, в основном компонентов клеточной стенки и капсулы. Наконец, в цитоплазматической мембране находятся важнейшие ферменты (биологические катализаторы). Упорядоченное расположение ферментов на мембранах позволяет регулировать их активность и предотвращать разрушение одних ферментов другими. С мембраной связаны рибосомы — структурные частицы, на которых синтезируется белок. Мембрана состоит из липопротеидов. Она достаточно прочна и может обеспечить временное существование клетки без оболочки. Цитоплазматическая мембрана составляет до 20% сухой массы клетки.
На электронных фотографиях тонких срезов бактерий цитоплазматическая мембрана представляется в виде непрерывного тяжа толщиной около 75А, состоящего из светлого слоя (липиды), заключенного между двумя более темными (белки). Каждый слой имеет ширину 20—30 А. Такая мембрана называется элементарной (табл. 30, рис. 8).
Между плазматической мембраной и клеточной стенкой имеется связь в виде десмозов -мостиков. Цитоплазматическая мембрана часто дает инвагинации — впячивания внутрь клетки. Эти впячивания образуют в цитоплазме особые мембранные структуры, названные мезосомами. Некоторые виды мезосом представляют собой тельца, отделенные от цитоплазмы собственной мембраной. Внутри таких мембранных мешочков упакованы многочисленные пузырьки и канальцы (рис. 2). Эти структуры выполняют у бактерий самые различные функции. Одни из этих структур — аналоги митохондрий. Другие выполняют функции эндо-плазматической сети или аппарата Гольджи. Путем инвагинации цитоплазматической мембраны образуется также фотосинтезирующий аппарат бактерий. После впячивания цитоплазмы мембрана продолжает расти и образует стопки (табл. 30), которые по аналогии с гранулами хлоропластов растений называют стопками ти-лакоидов. В этих мембранах, часто заполняющих собой большую часть цитоплазмы бактериальной клетки, локализуются пигменты (бактериохлорофилл, каротиноиды) и ферменты (цитохромы), осуществляющие процесс фотосинтеза.
В цитоплазме бактерий содержатся рибосомы— белок-синтезирующие частицы диаметром 200А. В клетке их насчитывается больше тысячи. Состоят рибосомы из РНК и белка. У бактерий многие рибосомы расположены в цитоплазме свободно, некоторые из них могут быть связаны с мембранами.
Рибосомы являются центрами синтеза белка в клетке. При этом они часто соединяются между собой, образуя агрегаты, называемые полирибосомами или полисомами.
В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Однако их присутствие нельзя рассматривать как какой-то постоянный признак микроорганизма, обычно оно в значительной степени связано с физическими и химическими условиями среды. Многие цитоплазматические включения состоят из соединений, которые служат источником энергии и углерода. Эти запасные вещества образуются, когда организм снабжается достаточным количеством питательных веществ, и, наоборот, используются, когда организм попадает в условия, менее благоприятные в отношении питания.
У многих бактерий гранулы состоят из крахмала или других полисахаридов -гликогена и гранулезы. У некоторых бактерий при выращивании на богатой сахарами среде внутри клетки встречаются капельки жира. Другим широко распространенным типом гранулярных включений является волютин (метахроматиновые гранулы). Эти гранулы состоят из полиметафосфата (запасное вещество, включающее остатки фосфорной кислоты). Полиметафосфат служит источником фосфатных групп и энергии для организма. Бактерии чаще накапливают волютин в необычных условиях питания, например на среде, не содержащей серы. В цитоплазме некоторых серных бактерий находятся капельки серы.
Помимо различных структурных компонентов, цитоплазма состоит из жидкой части — растворимой фракции. В ней содержатся белки, различные ферменты, т-РНК, некоторые пигменты и низкомолекулярные соединения — сахара, аминокислоты.
В результате наличия в цитоплазме низкомолекулярных соединений возникает разность в осмотическом давлении клеточного содержимого и наружной среды, причем у разных микроорганизмов это давление может быть различным. Наибольшее осмотическое давление отмечено у грамположительных бактерий — 30 атм, у грамотрицательных бактерий оно гораздо ниже — 4—8 атм.
Ядерный аппарат.
В центральной части клетки локализовано ядерное вещество — дезоксирибонуклеиновая кислота (ДНК).
У бактерий нет такого ядра, как у высших организмов (эукариотов), а есть его аналог — «ядерный эквивалент» - нуклеоид (см. рис. 2, 8), который является эволюционно более примитивной формой организации ядерного вещества. Микроорганизмы, не имеющие настоящего ядра, а обладающие его аналогом, относятся к прокариотам. Все бактерии -прокариоты. В клетках большинства бактерий основное количество ДНК сконцентрировано в одном или нескольких местах. В клетках эукариотов ДНК находится в определенной структуре — ядре. Ядро окружено оболочкой— мембраной.У бактерий ДНК упакована менее плотно, в отличие от истинных ядер; нуклеоид не обладает мембраной, ядрышком и набором хромосом. Бактериальная ДНК не связана с основными белками — гистонами — ив нуклео-иде расположена в виде пучка фибрилл.
Жгутики.
На поверхности некоторых бактерий имеются придаточные структуры; наиболее широко распространенными из них являются жгутики — органы движения бактерий.
Жгутик закрепляется под цитоплазматической мембраной с помощью двух пар дисков (см. на стр. 206). У бактерий может быть один, два или много жгутиков. Расположение их различно: на одном конце клетки, на двух, по всей поверхности и т. д. (рис. 9). Жгутики бактерий имеют диаметр 0,01—0,03 мкм, длина их может во много раз превосходить длину клетки. Бактериальные жгутики состоят из белка — флагеллина — и представляют собой скрученные винтообразные нити.
На поверхности некоторых бактериальных клеток имеются тонкие ворсинки — фимбрии. Подробнее они описаны на странице 205.
Обмен веществ у бактерии
При сравнительно бедных морфологических признаках бактерии отличаются большим разнообразием осуществляемых ими в природе превращений веществ.
Бактерии в совокупности с другими группами микроорганизмов выполняют колоссальную химическую работу. При их участии происходит разложение сложных органических веществ — растительных и животных отстатков — до простых минеральных соединений: углекислоты, аммиака, нитратов, сульфатов и др.,— которые вновь ассимилируются растениями, а затем поступают в организм животного. Таким образом на Земле осуществляется в колоссальном масштабе круговорот жизненно необходимых элементов: углерода, азота, серы, фосфора, железа и др., и бактерии являются важнейшим звеном в этом процессе.
Превращая различные соединения, бактерии получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий чрезвычайно разнообразны.
Одни из бактерий нуждаются в готовых органических веществах — аминокислотах, углеводах, витаминах,— которые должны присутствовать в среде, так как сами не могут их синтезировать. Такие микроорганизмы называются гетеротрофами. Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют исключительно за счет углекислоты. Они называются автотрофами.
По своим потребностям гетеротрофы очень разнообразны: некоторые из них нуждаются в большом наборе аминокислот, витаминов, углеводов и т. д.; другие требуют наличия в среде лишь небольшого числа готовых аминокислот, потребности в витаминах у них могут быть ограничены. Есть и такие формы, которые могут сами синтезировать все вещества: белки, сахара, жиры и т. д., если в среде, где происходит их развитие, присутствует всего одно или несколько простых органических соединений. Такие гетеротрофные организмы ближе стоят к автотрофам.
Каждый организм для поддержания жизни и осуществления процессов, совокупность которых составляет обмен веществ, нуждается в постоянном и непрерывном притоке энергии.
Гетеротрофные микроорганизмы получают энергию при окислении органических веществ кислородом или при сбраживании (без участия кислорода).
Типы окислительных процессов в мире бактерий исключительно разнообразны. Эти микроорганизмы могут окислять любые имеющиеся в природе органические вещества. Если бы в природе существовало какое-либо органическое вещество (продукт животного или растительного происхождения), которое не могло бы быть окислено каким-либо микробом, то оно неизбежно накапливалось бы на поверхности Земли, а этого не происходит. Только в недрах, изолированных от кислорода, могут сохраняться органические вещества — нефть, уголь. Против микробного окисления не могут устоять даже искусственно полученные синтетические вещества, отсутствующие в природе. Но не каждый вид бактерий может разлагать все органические вещества.
Есть формы, приспособленные к использованию лишь небольшого числа веществ, есть и более универсальные.
Более того, бактерии способны окислять не только органические, но и неорганические соединения. Окисление бактериями неорганических веществ — серы, аммиака, нитратов, соединений железа, водорода и др., в процессе которого происходит синтез органических веществ из углекислоты, называется хемосинтезом, а бактерии, осуществляющие этот процесс,— хемосинтетиками.
Различные вещества могут окисляться не только кислородом воздуха, но и соединениями, богатыми кислородом: нитратами, сульфатами и карбонатами. Денитрифицирующие и такие специализированные бактерии, как десульфати-рующие и метановые, в анаэробных условиях могут окислять органические, а также неорганические вещества при помощи этих соединений, которые при этом восстанавливаются соответственно до азота, аммиака, водорода и метана.
Особенностью окисления органических веществ бактериями, как и другими микробами, является то, что оно не обязательно идет до конца как дыхание, т. е. до образования углекислого газа и воды, и в среде остаются продукты неполного окисления.
Механизмы окислительных процессов у микроорганизмов часто включают те или иные стадии дыхания. Огромное разнообразие окисляемых веществ предполагает существование разных механизмов окисления.
Дыхание
Дыхание является самой совершенной формой окислительного процесса и наиболее эффективным способом получения энергии. Главное преимущество дыхания состоит в том, что энергия окисляемого вещества —• субстрата, на котором микроорганизм растет, используется наиболее полно. Поэтому в процессе дыхания перерабатывается гораздо меньше субстрата для получения определенного количества энергии, чем, например, при брожениях.
Рис.10. Роль пировнноградной кислоты в процессах дыхания и брожения.
Процесс дыхания заключается в том, что углеводы (или белки, жиры и другие запасные вещества клетки) разлагаются, окисляясь кислородом воздуха, до углекислого газа и воды. Выделяющаяся при этом энергия расходуется на поддержание жизнедеятельности организмов, рост и размножение. Бактерии вследствие ничтожно малых размеров своего тела не могут накапливать значительного количества запасных веществ. Поэтому они используют в основном питательные соединения среды.
В общем виде дыхание можно представить следующим уравнением:
С6Н12О6 + 602 = 6С02 + 6Н20 + 2,87-106 дж.
глюкоза кислород углекислый газ вода энергия
За этой простой формулой скрывается сложная цепь химических реакций, каждая из которых катализируется специфическим ферментом.
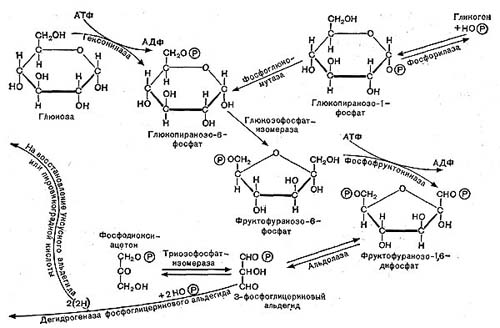
Рис. 11. Схема гликолитического пути расщепления углеводов.
Ферментативные реакции, происходящие в процессе дыхания, в настоящее время хорошо изучены. Схема реакций оказалась универсальной, т. е. в принципе одинаковой у животных, растений и многих микроорганизмов, в том числе бактерий. Процесс дыхания при окислении глюкозы складывается из следующих основных этапов (рис. 10).
Сначала происходит образование фосфорных эфиров глюкозы — моноaктивированная глюкоза в форме дифосфата далее расщепляется на два триозофосфата (трехуглеродные соединения): фосфоглицериновый альдегид и диоксиацетон-фосфат, которые могут обратимо превращаться друг в друга.
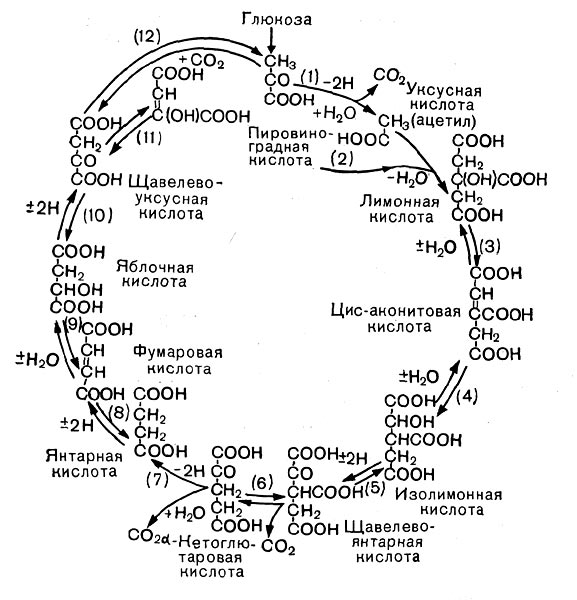
Рис. 12. Цикл трикарбоновых кислот. Стрелками показано направление, а номерами — порядок реакций.
Далее в обмен вступает фосфоглицериновый альдегид, он окисляется в дифосфо-глицериновую кислоту. Назначение этого процесса заключается в отщеплении атомов водорода от окисляемого субстрата и переносе водорода с помощью специфических окислительных ферментов к кислороду воздуха (см. рис. 10, 11).
Водород от фосфоглицеринового альдегида присоединяется к ферменту — никотин-амиддинуклеотиду (НАД); при этом альдегид окисляется до кислоты и выделяется энергия. Часть этой энергии тратится на образование АТФ; при этом присоединяется фосфорная кислота к аденозиндифосфат у— АДФ. При гидролизе АТФ энергия освобождается и может быть затрачена на различные процессы синтеза белка и другие нужды клетки.
Фосфоглицериыовая кислота окисляется до пировиноградной кислоты. При этом также образуется АТФ, т. е. запасается энергия.
На этом завершается первая — анаэробная — стадия процесса дыхания, которая носит название гликолитического пути или пути Эмбдена — Мейергофа — Парнаса. Для осуществления этих реакций кислород не требуется. Образовавшаяся пировиноградная кислота (СН3СОСООН) является интереснейшим и очень важным соединением. Пути расщепления глюкозы в процессе дыхания и многих брожений, вплоть до образования пировиноградной кислоты, идут совершенно одинаково, что впервые было установлено русским биохимиком С. П. Костычевым. Пировиноградная кислота является тем центральным пунктом, от которого расходятся пути дыхания и брожений, откуда начинается специфическая для данного процесса цепь ферментативных превращений -специфическая цепь химических реакций (рис. 11).
В процессе дыхания пировиноградная кислота вступает в цикл трикарбоновых кислот (рис. 12). Это сложный замкнутый круг превращений, в результате которых образуются органические кислоты с 4, 5 и 6 атомами углерода (яблочная, молочная, фумаро-вая, а-кетоглутаровая и лимонная) и отщепля-«тся углекислота.
Прежде всего от пировиноградной кислоты, содержащей три атома углерода, отщепляется СО2 — образуется уксусная кислота, которая с коферментом А образует активное соединение — ацетилкоэнзим А. Он передает остаток уксусной кислоты (ацетил) на щавелевоуксусную кислоту {4 атома углерода), и образуется лимонная кислота (6 атомов углерода). Лимонная кислота претерпевает несколько превращений, в результате выделяется С02 и образуется пяти-углеродное соединение — а-кетоглутаровая кислота. От нее тоже отщепляется С02 (третья молекула углекислого газа), и образуется янтарная кислота (4 атома углерода), которая затем превращается в фумаровую, яблочную и, наконец, щавелевоуксусную кислоту.
На этом цикл замыкается. Щавелевоуксусная кислота снова может вступить в цикл.
Таким образом, в цикл вступает трехуглерод-ная пировиноградная кислота, и по ходу превращений выделяются 3 молекулы С02.
Водород пировиноградной кислоты, освобождающийся при дегидрировании в аэробных условиях, не остается свободным — он поступает в дыхательную цепь (так же, как водород глицеринового альдегида, отнятый при превращении его в глицериновую кислоту). Это — цепь окислительных ферментов.
Ферменты, которые первыми берут на себя водород от окисляемого субстрата, называются первичными дегидрогеназами.
В их состав входят ди- или трипиридин-нуклео-тиды: НАД или НАДФ и специфический белок. Механизм присоединения водорода — один и тот же:
Окисляемое вещество — Н2 + НАД -> окисленное вещество + НАД*Н2
Водород, полученный дегидрогеназой, затем присоединяется к следующей ферментной системе — флавиновым ферментам (ФМН или ФАД).
От флавиновых ферментов электроны попадают на цитохромы — железосодержащие протеиды (сложные белки). По цепи цитохро-мов передается не атом водорода, а только электроны. При этом происходит изменение валентности железа: Fe++ — e->Fe++
Заключительная реакция дыхания — это присоединение протона и электрона к кислороду воздуха и образование воды. Но прежде происходит активирование молекулы кислорода под действием фермента цитохромоксидазы. Активирование сводится к тому, что кислород приобретает отрицательный заряд за счет присоединения электрона окисляемого вещества. К активированному кислороду присоединяется водород (протон), образуя воду.
Кроме упомянутой цепи переносчиков электронов и водорода, известны и другие. Процесс этот гораздо более сложен, чем изложенная схема.
Биологический смысл этих превращений заключается в окислении веществ и образовании энергии, В результате окисления молекулы сахара (глюкозы) в АТФ запасается 12,6-1053ж энергии, в самой молекуле сахара содержится 28,6-106 дж, следовательно, полезно используется 44% энергии. Это очень высокий коэффициент полезного действия, если сравнить его с к. п. д. современных машин.
В процессе дыхания образуется огромное количество энергии. Если вся она выделилась бы сразу, то клетка перестала бы существовать. Но этого не происходит, потому что энергия выделяется не вся сразу, а ступенчато, небольшими порциями. Выделение энергии небольшими дозами обусловлено тем, что дыхание представляет собой многоступенчатый процесс, на отдельных этапах которого образуются различные промежуточные продукты (с разной длиной углеродной цепочки) и выделяется энергия. Выделяющаяся энергия не расходуется в виде тепла, а запасается в универсальном макроэр-гическом соединении — АТФ. При расщеплении АТФ энергия может использоваться в любых процессах, необходимых для поддержания жизнедеятельности организма: на синтез различных органических веществ, механическую работу, поддержание осмотического давления протоплазмы и т. д.
Дыхание является процессом, дающим энергию, однако его биологическое значение этим не ограничивается. В результате химических реакций, сопровождающих дыхание, образуется большое количество промежуточных соединений. Из этих соединений, имеющих различное количество углеродных атомов, могут синтезироваться самые разнообразные вещества клетки: аминокислоты, жирные кислоты, жиры, белки, витамины.
Поэтому обмен углеводов определяет остальные обмены веществ (белков, жиров). В этом его огромное значение.
С процессом дыхания, его химическими реакциями связано одно из удивительных свойств микробов — способность испускать видимый свет — люминесцировать.
Известно, что ряд живых организмов, в том числе бактерии, могут испускать видимый свет. Люминесценция, вызываемая микроорганизмами, известна уже в течение столетий. Скопление люминесцирующих бактерий, находящихся в симбиозе с мелкими морскими животными, иногда приводит к свечению моря; с люминесценцией встречались также при росте некоторых бактерий на мясе и т. д.
К основным компонентам, взаимодействие между которыми приводит к испусканию света, относятся восстановленные формы ФМН или НАД, молекулярный кислород, фермент люцифераза и окисляемое соединение — люциферин. Предполагается, что восстановленные НАД или ФМН реагируют с люциферазой, кислородом и люциферином, в результате чего электроны в некоторых молекулах переходят в возбужденное состояние и возвращение этих электронов на основной уровень сопровождается испусканием света. Люминесценцию у микробов рассматривают как «расточительный процесс», так как при этом энергетическая эффективность дыхания снижается.
Брожение
Жизнь микробов возможна и без доступа кислорода воздуха. Энергия, необходимая для жизнедеятельности организма, в этих условиях образуется в результате процессов брожения. Наиболее распространены виды брожений, в процессе которых происходит распад органических веществ (преимущественно Сахаров) под влиянием микроорганизмов, представляющий совокупность окислительно-восстановительных реакций. Брожения никогда не приводят к полному окислению органических веществ. Многие характерные формы брожения протекают без участия кислорода воздуха — анаэробно.
Поскольку свободный кислород, имеющийся на нашей планете, образовался в результате фотосинтеза, возникшего на более поздних этапах развития жизни на Земле, совершенно очевидно, что анаэробный способ извлечения энергии — брожение — более древний, чем процесс дыхания.
Брожение известно людям с незапамятных времен. Тысячелетиями человек пользовался спиртовым брожением при изготовлении вина. Еще раньше было известно о молочнокислом брожении. Люди употребляли в пищу молочные продукты, готовили сыры. При этом они не подозревали, что эти процессы происходят с помощью микроорганизмов. Термин «брожение» был введен голландским алхимиком Ван Хельмонтом в XVII в. для процессов, идущих с выделением газов (fermentatio — кипение). Затем в XIX в. основоположник современной микробиологии Луи Пастер показал, что брожение является результатом жизнедеятельности микробов, и установил, что различные брожения вызываются разными микроорганизмами.
Спиртовое брожение
Спиртовое брожение — это процесс окисления углеводов, в результате которого образуются этиловый спирт, углекислота и выделяется энергия.Сбраживание cахаров известно с глубокой древности. В течение столетий пивовары и виноделы использовали способность некоторых дрожжей вызывать спиртовое брожение, в результате которого сахара превращаются в спирт.
Брожение производят главным образом дрожжи, а также некоторые бактерии и грибы. В различных странах для получения спирта используют различные микроорганизмы. Например, в Европе используют в основном дрожжи из рода Saccharomyces, в Южной Америке — бактерии Pseudomonas lindneri, в Азии — мукоровые грибы.
Сбраживаться могут лишь углеводы, и притом весьма избирательно. Дрожжи сбраживают только некоторые 6-углеродные сахара (глюкозу, фруктозу, маннозу).
Схематично спиртовое брожение может быть изображено уравнением
С6Н12О6 -> 2С2Н5ОН + 2С02 + 23,5 • 104 дж
глюкоза этиловый спирт углекислота энергия
Процесс спиртового брожения — многоступенчатый, состоящий из цепи химических реакций. Превращения глюкозы до образования пи-ровиноградной кислоты происходят так же, как и при дыхании. Эти реакции происходят без участия кислорода (анаэробно). Далее пути дыхания и брожения расходятся.
При спиртовом брожении пировиноградная кислота превращается в конечном итоге в спирт и углекислоту. Эти реакции протекают в две стадии. Сначала от пирувата отщепляется С02 и образуется уксусный альдегид; затем уксусный альдегид присоединяет водород, восстанавливаясь в этиловый спирт. Все реакции катализируются ферментами. В восстановлении альдегида участвует НАД-H2.
Обычно при спиртовом брожении, кроме главных продуктов, образуются побочные. Они довольно разнообразны, но присутствуют в небольшом количестве: амиловый, бутиловый и другие спирты, смесь которых называется сивушным маслом — соединение, от которого зависит специфический аромат вина. Образование побочных веществ связано с тем, что превращение глюкозы частично идет другими путями.
Биологический смысл спиртового брожения заключается в том, что образуется определенное количество энергии, которая запасается в форме АТФ, а затем расходуется на все жизненно необходимые процессы клетки.
Молочнокислое брожение.
При молочнокислом брожении конечным продуктом является молочная кислота.
С этим брожением люди знакомы издавна. Сквашивание молока, приготовление простокваши, кефира, квашение овощей — результаты молочнокислого сбраживания сахара молока или углеводов растений. Этот вид брожения осуществляется с помощью молочнокислых бактерий, которые подразделяются на две большие группы (в зависимости от характера брожения): гомоферментативные, образующие из сахара только молочную кислоту, и гетероферментативные, образующие, кроме молочной кислоты, спирт, уксусную кислоту, углекислый газ.
Гомоферментативное молочнокислое брожение вызывают бактерии рода Lactobacillus и стрептококки. Они могут сбраживать различные сахара с 6-ю (гексозы) или 5-ю (пентозы) углеродными атомами, некоторые кислоты. Однако круг сбраживаемых ими продуктов ограничен.
У молочнокислых бактерий нет ферментативного аппарата для использования кислорода воздуха. Кислород для них или безразличен, или угнетает развитие.
Молочнокислое брожение может быть описано уравнением
С6Н12О6 -> 2СН3*CНОН*СООН+21,8-104 дж
глюкоза молочная кислота энергия
Процесс образования молочной кислоты чрезвычайно близок к процессу спиртового брожения. Глюкоза также расщепляется до пирови-ноградной кислоты. Но затем ее декарбоксили-рование (отщепление С02), как при спиртовом брожении, не происходит, так как молочнокислые бактерии лишены соответствующих ферментов. У них активны дегидрогеназы (НАД). Поэтому пировиноградная кислота сама (а не уксусный альдегид, как при спиртовом брожении) принимает водород от восстановленной формы НАД и превращается в молочную кислоту. В процессе молочнокислого брожения бактерии получают энергию, необходимую им для развития в анаэробных условиях, где использование других источников энергии затруднено.
Гетероферментативное молочнокислое брожение — процесс более сложный, чем гомоферментативное: сбраживание углеводов приводит к образованию ряда соединений, накапливающихся в зависимости от условий процесса брожения. Одни бактерии образуют, помимо молочной кислоты, этиловый спирт и углекислоту, другие — уксусную кислоту; некоторые гетероферментативные молочнокислые бактерии могут образовывать различные спирты, глицерин, маннит.
Гетероферментативное молочнокислое брожение вызывают бактерии рода Lactobacterium и рода Streptococcus. Химизм этих брожений изучен не так хорошо, как спиртового или гомо-ферментативного молочнокислого брожения.
Гетероферментативные бактерии образуют молочную кислоту иным путем. Последняя стадия — восстановление пировиноградной кислоты до молочной — та же самая, что и в случае гомоферментативного брожения. Но сама пировиноградная кислота образуется при ином расщеплении глюкозы — гексозомонофосфат-ном. Выход энергии гораздо меньше, чем при спиртовом брожении.
Гетероферментативные бактерии сбраживают ограниченное число веществ: некоторые гексозы (причем определенного строения), пентозы, сахароспирты и кислоты.
Молочнокислое брожение широко используется при выработке молочных продуктов: простокваши, ацидофилина, творога, сметаны. При производстве кефира, кумыса наряду с молочнокислым брожением, вызываемым бактериями, имеет место и спиртовое брожение, вызываемое дрожжами. Молочнокислое брожение происходит на первом этапе изготовления сыра, затем молочнокислые бактерии сменяются пропионовокислыми.
Молочнокислые бактерии нашли широкое применение при консервировании плодов и овощей, в силосовании кормов. Чистое молочнокислое брожение применяется для получения молочной кислоты в промышленных масштабах.
Молочная кислота находит широкое применение в производстве кож, красильном деле, при выработке стиральных порошков, изготовлении пластмасс, в фармацевтической промышленности и во многих других отраслях. Молочная кислота также нужна в кондитерской промышленности и для приготовления безалкогольных напитков.
Маслянокислое брожение
Превращение углеводов с образованием масляной кислоты было известно давно. Природа маслянокислого брожения как результат жизнедеятельности микроорганизмов была установлена Луи Пасте-ром в 60-х годах прошлого века.
Возбудителями брожения являются масля-нокислые бактерии, получающие энергию для жизнедеятельности путем сбраживания углеводов. Они могут сбраживать разнообразные вещества — углеводы, спирты и кислоты, способны разлагать и сбраживать даже высокомолекулярные углеводы — крахмал, гликоген, декстрины.
Маслянокислое брожение в общем виде описывается уравнением
C6H12О6 -> СН3*CН2*СООН+2С02+2Н2
глюкоза масляная кислота
При этом брожении накапливаются различные побочные продукты. Наряду с масляной кислотой, углекислым газом и водородом образуются этиловый спирт, молочная и уксусная кислоты.
Некоторые маслянокислые бактерии, кроме того, образуют ацетон, бутанол и изопропи-ловый спирт.
Брожение начинается с процесса фосфорили-рования глюкозы и далее идет по гликолитиче-скому пути до стадии образования пировиноградной кислоты. Затем образуется уксусная кислота, которая активируется ферментом. После чего при конденсации (соединении) из двуугле-родного соединения получается четырехугле-родная масляная кислота. Таким образом, при маслянокислом брожении происходит не только разложение веществ, но и синтез.
По данным В. Н. Шапошникова, в маслянокислом брожении различаются две фазы. В первой параллельно с увеличением биомассы накапливается уксусная кислота, а масляная кислота образуется преимущественно во второй фазе, когда синтез веществ тела замедляется.
Маслянокислое брожение происходит в природных условиях в гигантских масштабах: на дне болот, в заболоченных почвах, илах и всех тех местах, куда ограничен доступ кислорода. Благодаря деятельности маслянокис-лых бактерий разлагаются огромные количества органического вещества.
Спиртовое, гомоферментативное молочнокислое и маслянокислое брожения являются основными типами брожений. Все другие виды брожений представляют собой комбинацию этих трех типов. Так, например, пропионовокислое брожение, играющее важную роль при производстве сыров и сопровождающееся накоплением пропионовой и уксусной кислот и углекислого газа, мо.жет рассматриваться как комбинация гомоферментативного молочнокислого и спиртового брожений. Брожения клетчатки и пектиновых веществ являются разновидностями маслянокислого брожения.
Итак, три основных типа брожения органически связаны между собой — начальные пути разложения углеводов у них одинаковы.
Процессы дыхания и брожения являются основными источниками энергии, необходимой микроорганизмам для нормальной жизнедеятельности, осуществления процессов синтеза важнейших органических соединений.
Биосинтез белка
В обмене веществ организма ведущая роль принадлежит белкам и нуклеиновым кислотам. Белковые вещества составляют основу всех жизненно важных структур клетки, они входят в состав цитоплазмы. Белки обладают необычайно высокой реакционной способностью. Они наделены каталитическими функциями, т. е. являются ферментами, поэтому белки определяют направление, скорость и теснейшую согласованность, сопряженность всех реакций обмена веществ.
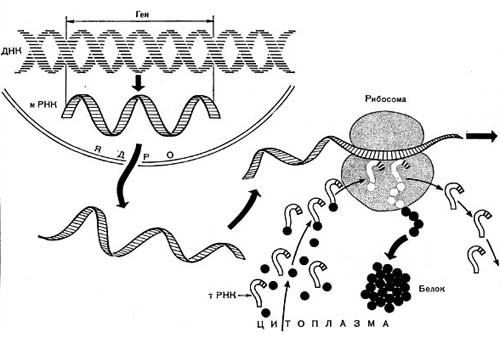
Рис. 13 А. Схема синтеза белка в эукариотной клетке.
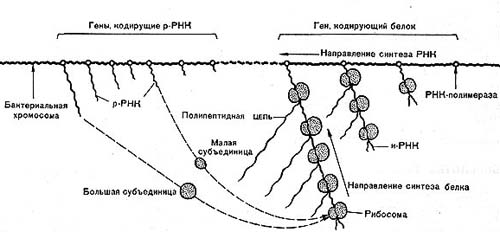
Рис. 13 Б. Схема синтеза белка в прокариотной клетке.
Ведущая роль белков в явлениях жизни связана с богатством и разнообразием их химических функций, с исключительной способностью к различным превращениям и взаимодействиям с другими простыми и сложными веществами, входящими в состав цитоплазмы.
Нуклеиновые кислоты входят в состав важнейшего органа клетки — ядра, а также цитоплазмы, рибосом, митохондрий и т. д. Нуклеиновые кислоты играют важную, первостепенную роль в наследственности, изменчивости организма, в синтезе белка.
Процесс синтеза белка является очень сложным многоступенчатым процессом. Совершаем ся он в специальных органеллах — рибосомах. В клетке содержится большое количество рибосом. Например, у кишечной палочки их около 20000.
Каким образом происходит синтез белка в рибосомах?
Молекулы белков по существу представляют собой полипептидные цепочки, составленные из отдельных аминокислот. Но аминокислоты недостаточно активны, чтобы соединиться между собой самостоятельно. Поэтому, прежде чем соединиться друг с другом и образовать молекулу белка, аминокислоты должны активироваться. Эта активация происходит под действием особых ферментов. Причем каждая аминокислота имеет свой, специфически настроенный на нее фермент.
Источником энергии для этого (как и для многих процессов в клетке) служит аденозинтрифосфат (АТФ).
В результате активирования аминокислота становится более лабильной и под действием того же фермента связывается с т-РНК.
Важным является то, что каждой аминокислоте соответствует строго специфическая т-РНК. Она находит «свою» аминокислоту и переносит ее в рибосому. Поэтому такая РНК и получила название транспортной.
Следовательно, в рибосому поступают различные активированные аминокислоты, соединенные со своими т-РНК. Рибосома представляет собой как бы конвейер для сборки цепочки белка из поступающих в него различных аминокислот (рис. 13 Аи Б).
Возникает вопрос: от чего зависит порядок связывания между собой отдельных аминокислот? Ведь именно этот порядок и определяет, какой белок будет синтезирован в рибосоме, так как от порядка расположения аминокислот в белке зависит его специфика. В клетке содержится более 2000 различных по строению и свойствам специфических белков.
Оказывается, что одновременно с т-РНК, на которой «сидит» своя аминокислота, в рибосому поступает «сигнал» от ДНК, которая содержится в ядре. В соответствии с этим сигналом в рибосоме синтезируется тот или иной белок, тот или иной фермент (так как ферменты являются белками).
Направляющее влияние ДНК на синтез белка осуществляется не непосредственно, а с помощью особого посредника, той формы РНК, которая получила название матричной или информационной РНК (м-РНК или и-РНК).
Информационная РНК синтезируется в ядре под влиянием ДНК, поэтому ее состав отражает состав ДНК. Молекула РНК представляет собой как бы слепок с формы ДНК.
Синтезированная и-РНК поступает в рибосому и как бы передает этой структуре план — в каком порядке должны соединяться друг с другом поступившие в рибосому активированные аминокислоты, чтобы синтезировался определенный белок. Иначе, генетическая информация, закодированная в ДНК, передается на и-РНК и далее на белок.
Молекула информационной РНК поступает в рибосому и как бы прошивает ее. Тот ее отрезок, который находится в данный момент в рибосоме, определенный ко доном (триплет), взаимодействует совершенно специфично с подходящим к нему по строению триплетом (антикодоном) в транспортной РНК, которая принесла в рибосому аминокислоту. Транспортная РНК со своей аминокислотой подходит к определенному кодону и-РНК и соединяется с ним; к следующему, соседнему участку и-РНК присоединяется другая т-РНК с другой аминокислотой и так далее, до тех пор пока не будет считана вся цепочка и-РНК и пока не нанижутся все аминокислоты в соответствующем порядке, образуя молекулу белка. А т-РНК, которая доставила аминокислоту к определенному участку полипептидной цепи, освобождается от своей аминокислоты и выходит из рибосомы. Затем снова в цитоплазме к ней может присоединиться нужная аминокислота, и она снова перенесет ее в рибосому. В процессе синтеза белка участвует одновременно не одна, а несколько рибосом — полирибосомы.
Основные этапы передачи генетической информации: синтез на ДНК как на матрице и-РНК (транскрипция) и синтез в рибосомах полипептидной цепи по программе, содержащейся в и-РНК (трансляция), универсальны для всех живых существ. Однако временные и пространственные взаимоотношения этих процессов различаются у про- и эукариотов.
У организмов, обладающих настоящим ядром (животные, растения), транскрипция и трансляция строго разделены в пространстве и времени: синтез различных РНК происходит в ядре, после чего молекулы РНК должны покинуть пределы ядра, пройдя через ядерную мембрану (рис. 13 А). Затем в цитоплазме РНК транспортируются к месту синтеза белка — рибосомам. Лишь после этого наступает следующий этап — трансляция.
У бактерий, ядерное вещество которых не отделено от цитоплазмы мембраной, транскрипция и трансляция идут одновременно (рис. 13 Б).
Современные схемы, иллюстрирующие работу генов, построены на основании логического анализа экспериментальных данных, полученных с помощью биохимических и генетических методов. Применение тонких электронно-микроскопических методов позволяет в буквальном смысле слова увидеть работу наследственного аппарата клетки. В последнее время получены электронно-микроскопические снимки, на которых видно, как на матрице бактериальной ДНК, в тех участках, где к ДНК прикреплены молекулы РНК-полимеразы (фермента, катализирующего транскрипцию ДНК в РНК), происходит синтез молекул и-РНК. Нити и-РНК, расположенные перпендикулярно к линейной молекуле ДНК, продвигаются вдоль матрицы и увеличиваются в длине. По мере удлинения нитей РНК к ним присоединяются рибосомы, которые, продвигаясь, в свою очередь, вдоль нити РНК по направлению к ДНК, ведут синтез белка.
Из всего сказанного следует, что местом синтеза белков и всех ферментов в клетке являются рибосомы. Образно выражаясь, это как бы «фабрики» белка, как бы сборочный цех, куда поступают все материалы, необходимые для сборки полипептидной цепочки белка из аминокислот. Природа же синтезируемого белка зависит от строения и-РНК, от порядка расположения в ней нуклеоидов, а строение и-РНК отражает строение ДНК, так что в конечном итоге специфическое строение белка, т. е. порядок расположения в нем различных аминокислот, зависит от порядка расположения нуклеоидов в ДНК, от строения ДНК.
Изложенная теория биосинтеза белка получила название матричной теории. Матричной эта теория называется потому, что нуклеиновые кислоты играют как бы роль матриц, в которых записана вся информация относительно последовательности аминокислотных остатков в молекуле белка.
Создание матричной теории биосинтеза белка и расшифровка аминокислотного кода является крупнейшим научным достижением XX века, важнейшим шагом на пути к выяснению молекулярного механизма наследственности.



