




Жизнь растений
Растение и среда
Жизнь растения, как и всякого другого живого организма, представляет сложную совокупность взаимосвязанных процессов; наиболее существенный из них, как известно, обмен веществ с окружающей средой. Среда является тем источником, откуда растение черпает пищевые материалы, затем перерабатывает их в своем теле, создавая такие же вещества, как те, из которых состоит тело растения, — совершается усвоение почерпнутых из среды веществ, их ассимиляция. Одновременно с этим процессом в организме совершается разрушение составных частей тела; разложение их на более простые. Этот противоположный процесс называют диссимиляцией. Ассимиляция, диссимиляция, неразрывно связанное с ними поступление веществ из окружающей среды и выделение в среду ненужных, отработанных — все это и есть обмен веществ. Следовательно, обменные явления тесно связывают организм растения со средой. Связь эта двоякая. Во-первых, растение оказывается зависимым от среды. В среде должны быть все необходимые для жизни растения материалы. Недостача, тем более отсутствие той или иной категории пищевых материалов должны привести к замедлению или даже прекращению жизненных явлений, к смерти. Во-вторых, поглощая из среды питательные вещества и выделяя в среду продукты своей жизнедеятельности (например, в форме опадающих листьев, омертвевших поверхностных слоев коры и т. п.), растение изменяет окружающую его среду. Следовательно, не только растение зависит от среды, но и среда всегда в какой-то мере зависит от растений.
Изменения среды растениями связаны не только с внесением в нее продуктов обмена веществ, но и с той физической работой, которую осуществляет растение. Когда корни растения внедряются в почву, они производят механическую работу разрушения или местного уплотнения субстрата. Работа, производимая растением, не ограничивается механическим воздействием на субстрат. В сущности, все физиологические функции растения представляют определенные формы работы. Это подводит к представлению о связях между растениями и средой и в ином плане: всякая работа связана с затратой энергии. Но энергия, как известно, «не исчезает и не творится вновь». Поэтому если растение расходует энергию, то, очевидно, оно должно откуда-то ее получать.
Источником энергии для растений, содержащих хлорофилл, служит лучистая энергия света, за счет которой растение строит органическое вещество, содержащее как бы законсервированную энергию. У растений, не имеющих хлорофилла, например грибов, источником энергии служит органическая пища, т. е. либо само созданное зеленым растением органическое вещество, либо оно же, но в форме, уже измененной другими организмами.
Энергия, в той или иной форме поступающая в растения, претерпевает в нем сложные изменения, выделяясь в конечном счете в окружающую среду. Можно сказать, что связь между растением и средой не ограничивается обменом и преобразованием веществ — параллельно этому совершается и энергетический обмен.
Среда жизни растения неоднородна, в составе ее можно отличить много компонентов, тесно связанных друг с другом. Каждый из элементов среды, оказывающий воздействие на организм, называют экологическим фактором. Разнообразие экологических факторов можно объединить в две категории: факторы биотические и факторы абиотические.
Факторы биотические — это организмы, окружающие данное растение. Они могут влиять на растение двояко. Очень часто мы встречаемся с тем, что организмы, окружающие данное растение, изменяют физическую среду жизни растения, влияя на ее температурный режим, на влажность воздуха, на освещенность и пр. Кроме такого опосредствованного воздействия, передаваемого через среду влияния, биотические факторы оказывают и непосредственное воздействие. Так, многие животные питаются определенными органами растений и целыми растениями (травоядные животные, паразиты из животного и растительного мира). В других случаях живые организмы оказываются более или менее полезными, способствуя оплодотворению растений (опылители), а также распространению семян. Биотические факторы обычно находятся в сложных взаимных связях.
Часто бывает так, что влияние, например, какого-нибудь животного на данное растение осуществляется через целую цепь живых звеньев. Птица, уничтожая насекомых-опылителей, может повлиять на возникновение и численность потомства данного растения.
Факторы абиотической группы, подобно биотическим, тоже находятся в определенных взаимодействиях. Например, при отсутствии воды элементы минерального питания, находящиеся в почве, становятся недоступными растениям; высокая концентрация солей в почвенном растворе затрудняет и ограничивает поглощение растением воды; ветер усиливает испарение и, следовательно, потерю растением воды; повышенная интенсивность света связана с повышением температуры среды и самого растения. Подобного рода связей известно много, иногда при ближайшем исследовании они оказываются очень сложными.
Изучая взаимоотношения между растениями и средой, нельзя противопоставлять биотические и абиотические компоненты среды, представлять эти компоненты самостоятельными, изолированными друг от друга; наоборот, они тесно связаны, как бы взаимопроникают друг в друга.
Так, пожизненные остатки всех растений (и животных), попадая в субстрат, изменяют его (влияние биотическое), привнося, например, элементы минерального питания, находившиеся в связанном состоянии в теле организмов; за счет этих элементов (влияние абиотическое) в какой-то степени возрастает плодородие субстрата, а это отражается на количестве растительной массы, т. е. в усилении биотического компонента среды (фактор биотический). Подобный простой пример показывает, что и биотические и абиотические факторы теснейшим образом переплетаются. Таким образом, окружение каждого растения рисуется как единство, как целостное явление, называемое средой.
Исследование среды и отдельных ее элементов составляет одну из важнейших задач ботаники. Сведения о роли каждого из факторов в жизни растения, в образовании им растительной массы и отдельных продуктов можно использовать для направленного воздействия на растения.
Но выявить значение отдельных факторов не всегда просто. Решение такой задачи облегчается при установлении некоторых общих закономерностей. Укажем две наиболее общие закономерности.
Первая носит название закона относительного действияфакторов и формулируется так: относительное действие фактора тем больше, чем больше этот фактор приближается к минимуму по сравнению с прочими. Относительное влияние фактора падает с повышением его интенсивности и приближается в области максимума к нулевому значению. Содержание этого закона сводится к следующему: если растение достаточно обеспечено каким-нибудь фактором, необходимым для жизни, например водой, то усиление водоснабжения хотя и может дать некоторый эффект, но этот эффект будет незначительным по сравнению с эффектом от увеличения напряженности фактора, который находится в состоянии, близком к минимуму.
Эта закономерность первоначально была установлена для обеспеченности растения элементами минерального питания, но позднее распространена и на другие, химические и физические, факторы.
Минимальное значение фактора — это такая напряженность фактора, которая приближается к пределу выносливости растения или переходит этот предел.
Закон относительного действия факторов позволяет путем наблюдения за растением в различных местообитаниях с известной вероятностью определить, напряженность каких факторов и в каких местах близка к минимуму и имеет ли она здесь решающее значение для растения. Точное же выявление факторов минимальной напряженности требует экологического и даже физиологического эксперимента.
Вторая закономерность, важная для понимания значения отдельных факторов для растения, состоит в том, что возрастание напряженности фактора, начиная с его предельно малого значения, вызывает сначала повышение эффекта, а затем, после достижения некоторой степени напряженности, дальнейшее ее нарастание оказывает отрицательное влияние, что приводит в итоге ко второму пределу выносливости растения. То же относится и к зависимости отдельных функций от факторов.
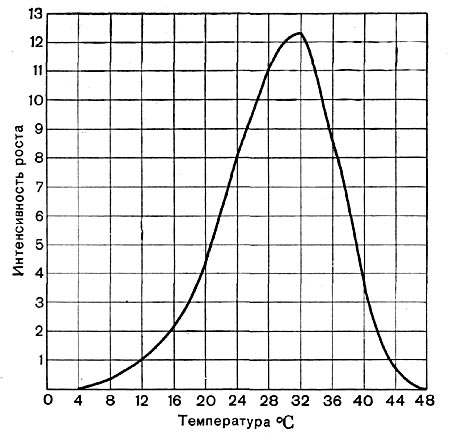
Рис.40. Рост проростков кукурузы при различных температурах.
Сущность этой закономерности сводится к тому, что по отношению к любому фактору существуют наименьшая (минимальная) и наибольшая (максимальная) его напряженности, начиная с которых и за пределами амплитуды, определяемой ими, существование организма (или существование отдельных его функций) невозможно. Между минимальной и максимальной напряженностями есть некоторая наилучшая (оптимальная) для растений (или для изучаемой функции его) напряженность, соответствующая наилучшему для растения состоянию или наибольшему эффекту рассматриваемой функции. Поясним это простым примером. Начиная с крайнего недостатка воды в почве, при котором, однако, растение может еще существовать, возрастание влажности почвы будет благоприятно отражаться на растении, но, начиная с некоторой достаточно высокой ступени оводненности почвы, дальнейшее, еще большее увлажнение неблагоприятно отразится на растении.
По отношению к отдельной функции организма эта закономерность вполне ясна. Так, на рисунке 40 показана интенсивность роста проростков кукурузы в зависимости от температуры. Наибольшая интенсивность роста в данном случае достигается при 30—32° С, а амплитуда температур, при которых возможен рост проростков, определяется минимальной температурой +4° и максимальной +48° С.
Изучение влияния напряженности факторов на отдельные функции растения показывает, что иногда оптимальная напряженность фактора не может быть оптимальной для организма растения в целом. Например, для оптимальной температуры дыхания выяснено, что по крайней мере в частных случаях она оказывается сублетальной (почти смертельной). Из этого следует, что оптимум отдельных функций не обязательно совпадает с оптимальной для организма в целом напряженностью фактора. Кроме того, оптимальные напряженности факторов часто не совпадают друг с другом. Например, оптимумы для фотосинтеза и дыхания обычно характеризуются различными температурами. Чем же определяется оптимальное состояние организма растения в целом? Однозначный, т. е. для всех растений применимый и всеми принимаемый, ответ на этот вопрос не получен. Однако во многих случаях можно воспользоваться показателями уклонений условий от оптимальных и таким образом очертить круг условий, приближающихся к оптимальным. Если в каких-то условиях наблюдается подавленность какой-нибудь важной сферы деятельности растения, то можно считать, что эта среда не обеспечивает растению наилучших условий существования. Так, например, отсутствие цветения или плодоношения, или резкое снижение (по сравнению с возможным) обилия цветков, плодов, семян, или угнетенный вид вегетативных органов, или устойчивое отсутствие проростков и молодых растений, или, наконец, снижение численности сравнительно с достижимой для данного растения в других условиях и тому подобное — все это можно считать выражением того, что в данном месте растение не находит себе наилучших условий. Определяя таким образом круг конкретных местообитаний, представляющих приближение условий к оптимальным, надо иметь в виду, что этот прием приводит к определению совокупности факторов (а не отдельного фактора), действующих взаимосвязанно на растение.
Выявлению влияния отдельных факторов способствуют некоторые географические данные.
Наблюдение за распространением растений показывает, что границы распространения иногда хорошо совпадают с некоторыми климатическими показателями. На карте 1 показана граница распространения одного из видов марены (Rubia peregrina) в Западной Европе. Эта граница хорошо совпадает с январской изотермой +4,5° С. В данном случае изотерма января, вероятно, определяет тот минимум среднемесячной температуры, при котором и выше которого может существовать данный вид марены. Еще более возможна обусловленность границы распространения в Европе вечнозеленого жестколистного кустарника падуба (Ilex aquifolium, карта 2). Его восточная граница проходит близко к январской изотерме 0° С и к линии, соединяющей пункты, в которых максимальная суточная температура выше 0° С наблюдается в течение по крайней мере 345 дней в году. В этом случае связь растения с тепловым фактором еще более вероятна.
Но в подобных случаях всегда все-таки остается сомнение в том, что именно данный фактор, а не какой-нибудь сопряженный с ним является прямой причиной ограничения области естественного распространения интересующего нас растения.
В таблице (справа) дано размещение коэффициента влажности на Восточно-Европейской равнине. Коэффициент влажности равен отношению количества осадков к показателю испарения. Таблица показывает, что к югу от южной границы ели, в лесостепной зоне, коэффициент влажности не превышает 15,0 и почти в 2/3 (64,5%) случаев характеризуется величинами 5,1—10,0. К северу от северной границы распространения ели во всех пунктах этот коэффициент более 25. На самой территории распространения ели в 77% пунктов коэффициент влажности изменяется в пределах от 10 де 20.
Из данных таблицы видно, что в пределах своего ареала ель преимущественно (77% точек) тяготеет к коэффициентам влажности от 10 до 20, а не к наиболее низким (5,1—10,0) и не к наиболее высоким (>25) значениям этой величины. Казалось бы, на основании этих данных можно было уверенно сказать, что граница ели и на севере и на юге определяется влажностью. В действительности такое категорическое заключение, по крайней мере относительно северной границы, неосторожно. Дело в том, что северная граница ели совпадает также с июльской изотермой +10° С. Очевидно, считать северную границу этой древесной породы, обусловленной только слишком высокой влажностью, нельзя; вероятно, существенным фактором является и температура. И нет ничего невероятного в том, что на севере высокие значения коэффициента влажности обусловливаются не столько количеством осадков, сколько низкими температурами и сами по себе, может быть, не создают препятствия расселению ели.
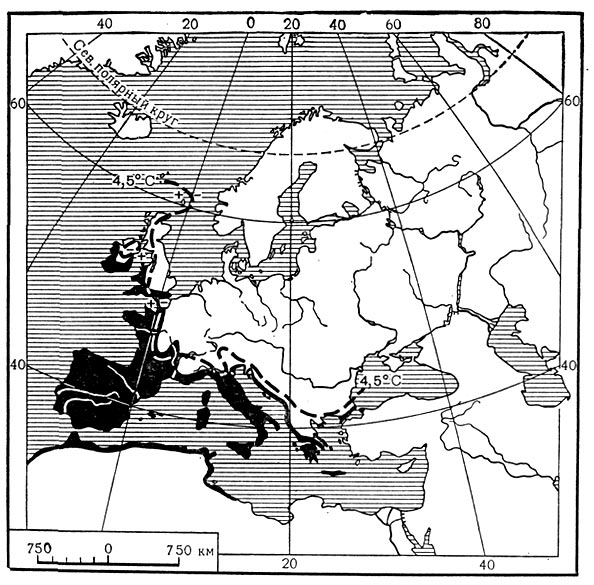
Карта 1. Распространение чужеземной марены в зависимости от температуры января.
Совпадение границ растений с так называемыми «вегетационными линиями» только в некоторой степени ориентируют наблюдателя в зависимости растения от отдельных факторов среды.
Немало дают и наблюдения за распределением интересующего исследователя вида в пределах ареала по разным местообитаниям, в особенности если условия заметно меняются на небольших расстояниях. Так, очень часто на склонах одни растения почти исключительно растут в верхней, другие в нижней части склона, а третьи занимают его середину. Нередко удается легко установить, что в этих местах меняются некоторые существенные жизненные условия: в частности, влажность почвы возрастает на склоне, как правило, по направлению сверху вниз. Исследователь принимает в качестве рабочей гипотезы следующее: растения, обитающие внизу склона, более влаголюбивы, чем растения, занимающие верхние участки, а наверху произрастают растения более суховыносливые. Затем дальнейшими наблюдениями исследователь подтверждает эту гипотезу или отказывается от сделанного предположения.

Коэффициент влажности на Восточно-Европейской равнине в пределах ареала ели и смежных с ним территорий
Наконец, в случае необходимости отношение растения к тому или иному фактору среды можно точно определить экспериментально.
Отношение различных растений к отдельным факторам и к их совокупности, как видно из приведенных примеров, коррелятивно связано с размещением растений по поверхности Земли как в широком географическом масштабе, так и в малом — топографическом.
Поэтому, находя растение, отношение которого к некоторым факторам уже выяснено, по присутствию этого растения, по состоянию его особей и по их численности можно достаточно уверенно судить о растительных условиях данного места.
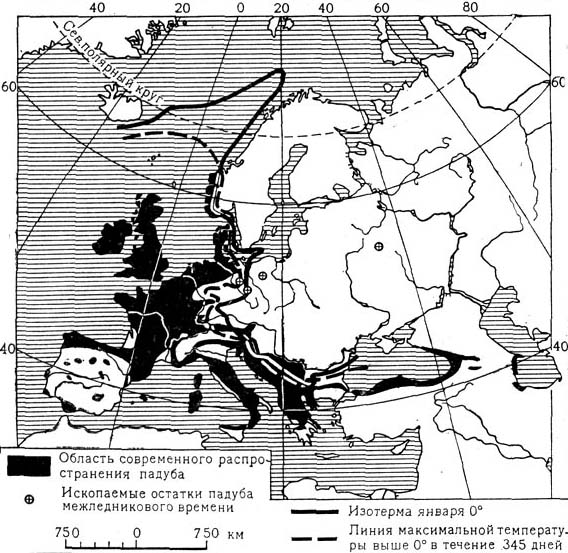
Карта 2. Зависимость распространения остролистного падуба о некоторых температурных условий.
Подобного рода сведения сначала (главным образом в XIX в.) имели сугубо частный характер и относились к растениям, связанным, например, с наличием в почве тех или иных химических элементов и их соединений в необычно большом количестве. Такие растения позднее стали называть индикаторами. Индикационные свойства растений стали использовать более широко, например для характеристики местообитаний по степени их влажности, минерального богатства и т. д., разделяя каждый такой признак местообитания на большое число градаций: от низкого увлажнения к высокому, от малого содержания солей к большому и т. п.
Растения-индикаторы и целые сообщества растений оказались пригодными для констатации содержания некоторых ценных ископаемых не только в почве, но и глубоко под ее поверхностью, что, в свою очередь, привело к возможности использования экологических данных в далекой от ботаники области — в геологических и гидрологических исследованиях при составлениицитологических карт, при установлении глубины залегания и химического состава грунтовых вод, при выявлении учабтков со значительной битуминозностью грунта и почвы. С помощью растений удается даже улавливать и фиксировать на картах линии тектонических нарушений.
Абиотические факторы делят на три группы — климатические, эдафические (почвенно-грун-товые) и орографические (связанные со строением земной поверхности). Первые две группы объединяют факторы, непосредственно своим влиянием определяющие те или иные стороны жизнедеятельности растения. Орографические факторы в основном выступают в роли видоизменяющих влияние прямодействующих.
Среди климатических факторов важное место в жизни растений занимают свет и тепло, связанные с лучистой энергией солнца; вода; состав и движение воздуха. Атмосферное давление и еще некоторые явления, входящие в понятие климата, существенного значения в жизни и распределении растений не имеют.
Свет и тепло поступают на Землю от Солнца. Энергетический поток, проходя через атмосферу, ослабляется, причем наиболее ослабевает ультрафиолетовый участок спектра. Ослабление потока солнечной энергии зависит от толщи атмосферы, которую проходят солнечные лучи, а следовательно, от географической широты, сезона и времени суток. Очень важно иметь в виду, что количество энергии, получаемое единицей земной поверхности, зависит от угла наклона поверхности, воспринимающей энергетический поток. Расчеты показывают, что на широте Ленинграда (60° с. ш.) южный склон крутизной 20° получает несколько большее количество солнечной радиации, чем горизонтальная поверхность на широте Харькова (50° с. ш.). В то же время на широте Харькова северный склон, имеющий крутизну 10°, получает меньшее количество солнечной радиации, чем горизонтальная поверхность на широте Ленинграда.
Поток энергии, достигающий твердой и водной оболочек Земли (литосферы и гидросферы), качественно отличен от того, который вступает в верхние разреженные слои атмосферы. От всей ультрафиолетовой радиации на земную поверхность попадают только сотые и тысячные доли калорий на 1 см2 в минуту, и здесь вовсе не обнаруживаются лучи с длиной волны 2800—2900 А, в то время как на высоте 50— 100 км ультрафиолетовая радиация содержит еще весь диапазон волн, включая и самые короткие.
Лучи с длиной волны от 3200 до 7800 А, охватывающие видимую (человеком) часть спектра, составляют лишь небольшую часть потока солнечной энергии, достигшей поверхности Земли.
Свет. Значение света определяется его участием в питании хлорофиллсодержащих растений. Свет поглощается хлорофиллом и используется на построение первичного органического вещества (фотосинтез). Кроме того, свет оказывает некоторое формообразующее воздействие на растения. Этому фактору принадлежит также существенная роль в достижении растениями цветения и плодоношения. В некоторых случаях свет оказывается существенным фактором прорастания семян.

Таблица 13. Альпийские растения: вверху (слева направо) — колокольчик, астра альпийская, фиалка; внизу слева — эдельвейс; внизу справа — мак альпийский.
Не весь поток солнечной энергии, падающей на зеленые части растения, усваивается ими. Некоторая часть лучей пропускается через листья, другая отражается. Эффект фотосинтеза зависит от силы света и выражается характерной кривой (рис. 41), показывающей, что одинаковые прибавки освещенности вызывают разное приращение эффекта фотосинтеза: большее при малом освещении и малое при большом. Разные растения неодинаково реагируют на изменение освещенности. В растениях, обычно обитающих в тени, фотосинтез быстрее достигает той интенсивности, после которой дальнейшее повышение освещенности почти не отражается на результатах фотосинтеза. Наоборот, светолюбивое растение с увеличением освещенности продолжает, хотя и медленно, наращивать усвоение углекислого газа.

Таблица 14. Жизненные формы (кустарники): вверху слева — «шпалерная» ива (Salix reticulata); вверху справа - толокнянка (Arctous alpma); внизу — кедровый стланик.
Общее количество света убывает от низких географических широт к высоким, но в том же направлении возрастает освещенная часть суток, а также доля рассеянного света, преимущественно используемого в фотосинтезе. В результате, принимая во внимание, что коэффициент использования света невелик (1—3%), разница в световых ресурсах средних и высоких широт незначительна. Подсчеты показывают, что световые условия фотосинтеза на широте Шпицбергена достаточны, чтобы растения могли производить столько же растительной массы, что и в средней части СССР. Малая продуктивность северной растительности объясняется не недостатком света, а недостатком тепла.
Повышенное содержание рассеянной радиации в высоких широтах объясняется высокой влажностью воздуха. Но такое положение существует и в других местах, в частности в приэкваториальной полосе. Здесь не раз отмечалась более низкая (общая) радиация, чем в умеренных широтах, что, вероятно, следует отнести за счет высокого содержания водяных паров. Таким образом, и в приэкваториальной полосе доля рассеянной радиации велика, но здесь она сочетается с благоприятными тепловыми условиями.
Значительные различия в условиях освещения наблюдаются между нижними и верхними поясами гор. В высокогорьях (по сравнению с соседними равнинами) общая радиация повышается в 2 раза и более, но это относится главным образом к прямому свету; рассеянное освещение остается на том же уровне или даже на более низком, чем на равнине. Кроме того, на горах освещенность возрастает еще за счет отражения света вблизи снега; поэтому альпийские растения, если они способны фотосинтези-ровать при низких температурах, оказываются в благоприятных световых условиях. В условиях высокогорий радиация богата ультрафиолетовыми лучами, влияние которых вообще вредно: по данным опытов, производимых в специальных камерах, под влиянием лучей с длиной волны 2800—3200 А большинство растений гибнет, кроме некоторых высокоальпийских, которые, однако, также испытывают угнетение в этих условиях (табл. 13).

Рис.41. Кривая усвоения СО2 световыми и теневыми растениями.-
Не менее специфичные световые условия создаются в тени. Будет ли затенение создано кронами деревьев или густым травостоем, под сомкнутым пологом более высоких растений светоснабжение всегда пониженное вследствие отражения и поглощения части светового потока совокупностью более высоких растений. Поэтому у самой поверхности почвы освещение всегда бывает более слабым, чем на поверхности растительности. Здесь, в затенении, если и поселяются какие-нибудь автотрофные растения, то только такие, которые могут довольствоваться ничтожными порциями световой энергии. В данном случае абиотический фактор (свет) преобразуется вследствие использования его более высокими растениями. Под пологом высоких растений свет не только ослабевает, во и качественно изменяется. Специальные исследования показали, что внутри леса световой спектр имеет два максимума — в красных (7100 А) и второй, меньший, в зеленых (5500 А) лучах. В лесу, следовательно, имеется «красно-зеленая тенистость». Почти весь свет в лесу рассеянный; прямое излучение проникает в виде неустойчивых бликов, меняющих свое положение в зависимости от высоты стояния солнца, сомкнутости крон, степени подвижности воздуха.
Из этого следует, что в лесу создается своеобразный световой климат, отличный от такового на открытых местах. Исследование поглощения света теневыми листьями показало, что кривые поглощения энергии соответствуют распределению ее в лесу.
Заметное ослабление силы света наблюдается также и в водной среде. Растения, погруженные в воду, никогда не получают свет, соответствующий его напряженности на соседнем, открытом месте. В зависимости от прозрачности воды и других обстоятельств убывание силы света в воде может идти в разном темпе, что в большой степени зависит от степени прозрачности (мутности) воды. Можно для примера указать, что в Средиземном море на глубине 1 м освещенность убывает до 82%; до глубины 5 м доходит свет, ослабленный более чем вдвое (45%); на глубине 30 м этот показатель сокращается еще в 10 раз (4,5%); на глубине 120 м освещенность составляет только 0,5% (1/200) от освещенности на поверхности. Эти данные сопоставимы с данными освещенности тенистых лесов. Так, кислица в еловых лесах может жить при освещенности, равной 1/200 от полного солнечного освещения, а в дождевых тропических лесах Суматры находили нормально растущие растения при освещенности, равной 1/500 и даже (споровые растения) при 1/1000 и 1/2ооо от полной. Ослабление освещения на глубине ставит водные растения в положение, аналогичное положению растений леса. Следовательно, погруженные водные растения — растения теневые.
Но световая обстановка их жизни отлична от «красно-зеленой» в тенистых лесах. В воде тенистость «зелено-голубая».
Изменение цветности освещения с глубиной в известной степени отражается на окраске глубинных водорослей. Некоторым из них свойственно усиление интенсивности зеленой окраски (увеличение содержания хлорофилла); другим же помогает окраска, дополнительная к цветности освещения, например окраска красных водорослей.
Надо упомянуть также и особенности освещения в пещерах, в глубоких расщелинах и т. д., во внутренние части которых не могут проникать прямые лучи солнечного света. Освещенность здесь может быть крайне низкой, составляя иногда ничтожную долю освещенности рядом находящихся открытых мест. Иногда внутри пещер освещенность составляет всего 1/1000 — 1/2500 от полного солнечного освещения. Тем не менее и при этих условиях обнаружены хлорофиллсодержащие растения! Проникать в столь затененные места, очевидно, могут далеко не все растения. Наблюдается следующее: способность проникновения растений в практически неосвещаемые глубины пещер, трещин и т. п. убывает в порядке водоросли -мхи — папоротники — семенные растения. Возможно, это отчасти объясняется тем, что водоросли, предростки мхов, иногда сами мхи и заростки папоротников практически состоят только из клеток, содержащих хлорофилл. Им не приходится расходовать продукты фотосинтеза на построение бесцветных частей тела и дыхание. Но, кроме того, у таких «сумеречных» растений должна быть особая способность к усвоению света столь ничтожной напряженности.
Нельзя не отметить и то, что свет малой напряженности может проникать в песчаные и трещиноватые почвы до глубины в несколько сантиметров, что позволяет поселяющимся здесь водорослям существовать за счет фотосинтеза. На влажных песчаных морских побережьях и пустошах на глубине нескольких миллиметров от поверхности можно найти слой сине-зеленых водорослей, которые своими слизистыми покровами склеивают песчинки, образуя тонкую, но плотную пленку. Можно было бы привести еще немало примеров подобного рода.
Разнообразие световых условий усугубляется еще и тем, что использование света как источника энергии в одних географических условиях возможно круглый год, в других ограничивается более или менее краткими сезонами.
Столь разнообразная световая среда, очевидно, может быть использована растениями, различно относящимися к свету.
Для приблизительной ориентировки в разнообразии растений по их отношению к свету, в основном к освещенности, пользуются показателем светового довольствия. Эта величина представляет отношение освещенности в месте, где живет растение, к освещенности на открытом (незатененном) месте. Пользуясь численной характеристикой светового довольствия, можно определить для каждого растения среднее (аср), минимальное (amin) и максимальное (a max) значения светового довольствия, т. е. определить световую амплитуду растения.
На основании таких данных различают следующие три категории растений:
1. Виды, которые почти исключительно живут на незатененных местах. аср =1; световая амплитуда растений очень мала, так как атах почти не отличается от amin. Таковы растения пустынь, степей, тундр, безлесных горных вершин и т. п. Это гелиофиты (от греч. «гелиос» — солнце, «фитон» — растение).
2. Виды, которые могут жить при полном солнечном освещении, но переносят и некоторое затенение. У них всегда имеется различие между максимальным и минимальным значениями светового довольствия. атах = 1, amin различно для разных видов. Отношение этих растений к освещенности может различаться в зависимости от возраста. В молодом, нецветущем состоянии растения часто способны переносить большее затенение, чем в возрасте плодоношения. К этой группе относятся многие растения, обычно произрастающие на лугах, ряд опушечных и даже некоторые типичные степные растения. Растения этой группы называют факультативными гелиофитами.
3. Виды, которые в природе никогда не растут на открытых местах; amax< 1, amjn всегда невелико, как правило, меньше, чем у факультативных гелиофитов. Примерами могут служить ветреница дубравная, яснотка крапчатая, сочевичник весенний и другие лесные растения. Растения этой группы называют скиофитами (от греч. «скиа» — тень) или сциофитами.
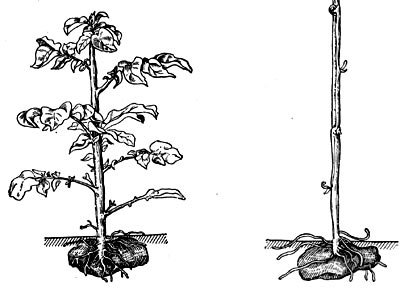
Рис.42. Этиолированный (справа) и нормальный (слева) побеги картофеля.
Эти три категории растений, конечно, нерезко разграничены. Кроме того, не всегда произрастание растения в освещенных (или затененных) местах свидетельствует об их действительной потребности в свете. Бывает, что растение довольствуется менее подходящими условиями из-за их вытеснения конкурентами с мест, более благоприятных для них по освещенности. Наконец, сама потребность в свете может измениться у одного и того же растения в разной климатической обстановке или при разных почвенных условиях.
Тем не менее в целом выделение этих трех групп вполне оправдано, так как по крайней мере два из них — гелиофиты и скиофиты — характеризуются достаточно определенными различиями. Если гелиофиты, или световые растения, слабо переносят затенение, то скиофиты, или теневые, переносят его хорошо. Световые растения достигают наибольшей интенсивности фотосинтеза при полном освещении, в то время как у теневых предельная интенсивность фотосинтеза отмечается при малой освещенности (рис. 41). Различаются эти растения и по интенсивности дыхания — она выше у световых и значительно меньше у теневых растений.
Наконец, как известно, в процессе фотосинтеза растения создают углеводы, на что потребляется углекислый газ; в процессе дыхания углеводы разрушаются с выделением углекислого газа. Интенсивность фотосинтеза зависит от напряженности света. При некоторой достаточно низкой напряженности света между фотосинтезом и дыханием создается такое соотношение, когда количество углекислого газа, выделяемого при дыхании, сравнимо с количеством, потребляемым при фотосинтезе. Такую силу света называют компенсационной точкой (пунктом). У световых растений компенсационный пункт соответствует более высокой освещенности, у теневых — слабой. Находясь под влиянием света разной напряженности и разного состава, растения несут как бы отпечаток этого в своем строении.
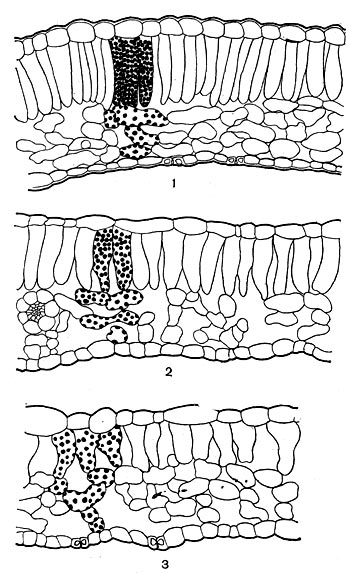
Рис. 43. Разрезы листьев лесной герани: 1 — солнечный лист; 2 и 3 — теневые листья.
Растения, обычно живущие при достаточной освещенности, попадая в затененные места, развивают этиолированные побеги (рис. 42). Это хорошо видно, например, на побегах картофеля, развившихся в темноте: они имеют белую, слегка желтоватую окраску; состоят из рыхлой, с большими межклетниками ткани, клетки которой вытянуты в длину; развитие листьев на таких побегах подавлено. Этиоля-ция рассматривается как результант влияния инфракрасных лучей (7300—8000 А), которые при недостатке более коротких лучей способствуют затягиванию второй фазы онтогенеза клеток — фазы растяжения. На свету эта фаза заканчивается быстрее.
Коротковолновые (синие и фиолетовые) лучи спектра стимулируют клеточные деления, но задерживают вторую фазу онтогенеза клетки — фазу растяжения.
Вероятно, именно этим объясняется приземистость травянистых растений, живущих на свету. С этим же, вероятно, связана низкорослость альпийских растений (табл. 13), живущих при повышенном количестве коротковолновой радиации. Приземистость, вызываемая свойствами света, оказывается биологически выгодной в альпийских условиях: она помогает использовать защитные свойства снега зимой.
Освещенность отражается и на различии в анатомическом строении листьев световых и теневых растений (рис. 43). Листья световых растений имеют хорошо выраженную палисадную ткань, а если растение живет на почве, отражающей много света (мел, известняк), то палисадная ткань может быть развита с обеих сторон листа. У световых растений эпидермис состоит из относительно мелких тонкостенных клеток, количество устьиц относительно велико. Скиофиты представляют противоположность гелиофитам и по этим признакам. Однако подобные различия связаны не столько с влиянием света, сколько с тем, что, живя на свету, скиофиты сильно нагреваются и это очень отражается на их водном режиме — световые растения часто являются в то же время растениями засушливых мест — ксерофитами.
Интересно отметить, что морфолого-анатоми-ческие особенности, свойственные световым и соответственно теневым растениям, могут проявляться и на одном и том же растении. Так, например, листья одного и того же дерева, взятые с незатененных ветвей и развившиеся в затенении, обычно хорошо различаются по анатомической структуре.
До сих пор мы рассматривали свет как фактор, имеющий непосредственное значение для фотосинтеза. Было упомянуто, что свету принадлежит известная роль в обеспечении достижения растением возможности производить цветки, осуществлять плодоношение. В этом отношении существенное влияние оказывает периодичность освещения.
Еще в начале 20-х годов XX в. было установлено, что сроки цветения многих растений можно сдвигать, изменяя соотношение между освещенной и неосвещенной частями суток. С этой точки зрения различают три группы растений. У растений длинного дня, поставленных в условия кратковременного ежесуточного освещения, наступление цветения задерживается и его может даже совсем не быть. В противоположность этому растения короткого дня ускоряют наступление цветения при кратком периоде ежесуточного освещения, и, наоборот, цветение у них запаздывает или вовсе не осуществляется при длинном дне. Третью группу составляют растения, у которых изменения длины дня не вызывают сдвигов во времени цветения.
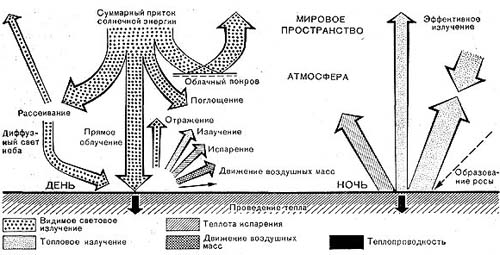
Рис.44. Приток и расход лучистой энергии.
Различное отношение к длине дня растений двух первых групп отражается на их географическом распространении и, возможно, связано с местом их возникновения. Если растение возникло в низких широтах, где день относительно короток, то такой вид должен был приспособиться к этому световому режиму. При расселении в область высоких широт такое растение может встретить препятствие в виде несоответствующей его потребностям большой продолжительности освещенной части суток. Не достигая плодоношения, оно окажется лишенным возможности дальнейшего распространения. Аналогичное можно сказать и о длинно-дневных растениях.
Отметим и то, что от освещения зависит иногда прорастание семян. У многих растений семена прорастают только в темноте. Это значит, что в природной обстановке семя должно обязательно оказаться в почве или под покровом подстилки (полуразложившиеся листья и другой опад). Надо ожидать, что в подобных случаях семена «пользуются услугами», например, животных, которые вовлекают семена в почву или под слой подстилки, или же «должны дожидаться» того времени, когда над отделившимся от материнского растения семенем накопится достаточно толстый и плотный слой спада. Например, семена Nigella sativa на свету не прорастают, в темноте же стопроцентное прорастание наступает уже через несколько дней. А семена Elatine alsinastrum годами могут лежать в темноте не прорастая, а на свету полное прорастание происходит в течение 2,5 недели. Более детальное исследование поведения семян вербейника позволило установить, что под действием облучения в 200 м/свеч в течение 5 мин прорастает более 40% семян, а для того, чтобы добиться этого результата при освещении 5 м/свеч, нужно облучать их около 3 ч. В некоторых случаях установлено, что для прорастания семян важна не только интенсивность света, но и его определенный качественный состав.
Тепло. Существенное значение тепла в жизни растений очевидно. Хорошо известно, что многие растения, вегетирующие круглый год в низких широтах, не переносят холодной зимы высоких широт. Никому не удавалось вырастить пальму в грунтовых условиях, на открытом месте где-нибудь под Москвой или Ленинградом. Известно, что лиственные деревья на зиму сбрасывают листву. В холодное, зимнее время практически прекращается рост побегов и корней, только у некоторых зимне-зеленых трав в конце зимы обнаруживается подснежный рост. В те месяцы года, когда температура держится на уровне ниже 0° С, видимые проявления жизни растений или вовсе прекращаются, или испытывают глубокую депрессию. В областях, где летняя температура очень высока, переход растения в состояние покоя, внешне сходное с зимним, приходится на время засухи. Выше были описаны примеры совпадения границ географического распространения отдельных растений с изотермами, что, вероятно, хотя бы отчасти объясняется температурным фактором. Все наблюдения подобного рода приводят к естественному заключению о большой важности теплового фактора в жизни растений.
Источник тепла — энергия солнечных лучей, которая в поглощенном виде может превратиться в тепловую. Это относится и к видимым и к не воспринимаемым нашим глазом инфракрасным лучам. Падая на поверхность почвы, энергетический поток частью отражается, частью поглощается ею, причем светлоокрашенные субстраты отражают больше лучей, темные — меньше. Поглощение осуществляется поверхностным слоем ничтожной мощности, что и приводит к его нагреванию. Тепло нагретого слоя переходит в глубже лежащие слои субстрата и в прилежащие к нему слои воздуха; оно расходуется на испарение (если в субстрате есть вода) и, наконец, излучается в атмосферу, главным образом в ясные ночи (рис. 44).
Энергетический поток поглощается не только почвой. В этом процессе участвуют и надземные части растения, причем большая часть энергии, поглощенной листьями (вообще хлорофиллсодержащими органами), у наземных растений тратится на испарение. Уже эти немногочисленные данные позволяют составить некоторое представление о разнообразии -тепловых условий, в которых живут растения.
Тепловая обстановка на суше определяется географическим положением (географической широтой и удаленностью от океана), рельефом (крутизна и направление склона, в горах — высота над уровнем моря), сезоном и, наконец, временем дня. Важным элементом теплового фона жизни является степень контрастности температуры, перепады ее от лета к зиме и от освещенной части суток к ночному времени.
Достаточно разнообразная, но гораздо более мягкая тепловая обстановка — в водоемах, особенно морских и океанических. Если понимать растительную жизнь как совокупность всего разнообразия растений, то, по-видимому, на Земле мало найдется таких мест, где по температурным условиям растительная жизнь вовсе исключалась бы. Она невозможна в кратерах действующих вулканов, но горячие ключи, распространенные в вулканических областях, населены автотрофными водорослями. Наиболее выдающимся примером могут служить сине-зеленые водоросли, которые существуют даже при +85,2° С в Северной Америке и при +77° С в горячих ключах на Камчатке. Это тем более удивительно, что белки протоплазмы обычно свертываются при температуре около +70° С. Того же порядка температуру могут переносить иногда и бактерии (сенная палочка и некоторые другие). В экспериментальных условиях установлено, что клетки дрожжевых грибков не теряют жизнеспособности при +90° С. Еще более жароустойчивы споры некоторых бактерий и грибов.
Надо иметь в виду, что и некоторые высшие наземные растения проводят часть своей жизни при достаточно высокой температуре. Так, у многих растений пустынь в полдень перегрев листьев достигает иногда 10—12° С по сравнению с окружающим воздухом. Повышение температуры тем значительнее, чем слабее испарение. У части растений, обитающих в местах, богатых водой, температура листьев на 3—6° С и более ниже окружающего воздуха. В общем же максимальная температура листьев даже у сильно испаряющих влагу растений превышает 40° С, а у лишенных возможности сильно испарять влагу может достигать 50° С. Одним из выдающихся в этом отношении растений является финиковая пальма, температура листьев которой достигает 53,5° С. Но даже столь высокая температура еще на 3—12° С ниже того предела, при котором наступает смерть. Виды растений крайне различны по отношению к избыточной температуре. Способность переносить подобную температуру связана частично со строением растений (мелкие листья, их слабая окраска, отражение лучей густым опушением, блестящей кутикулой, положение листьев по отношению к полуденным лучам и пр.), отчасти с внутренними особенностями, т. е. со свойствами протоплазмы.
В ряде случаев у низших гетеротрофных растений установлена способность переносить крайне низкие температуры, не наблюдаемые в природной обстановке. Например, грибница некоторых видов грибов из рода Мисог сохраняла жизнеспособность после охлаждения до — 110° С, а споры некоторых плесневых грибов из родов Penicillium и Aspergillus не теряли способность к прорастанию после охлаждения ниже —200° С.
Что же касается природных условий, то в Верхоянске, который до недавнего времени считался полюсом холода и где абсолютные минимумы температуры достигают — 68° С, развита достаточно богатая растительность лиственничных северотаежных лесов. Не менее замечательно, что на поверхности снегов высокогорий и высокоарктических широт могут жить некоторые водоросли. Чаще всего они бывают окрашены в красный цвет благодаря наличию красного пигмента — гематохрома, вследствие чего поверхность снега окрашивается в красный цвет («красный снег»). В других случаях окраска снега может быть зеленой, желтой, бурой (зеленые, сине-зеленые, диатомовые водоросли). Активная жизнь, в том числе и размножение этих организмов, обнаруживается при температуре, близкой к 0° С, когда поверхность снега оттаивает, чему способствует окраска водорослей, поглощающих солнечные лучи. Разнообразие таких нивальных (снежных) водорослей невелико и для высокогорий исчисляется всего несколькими десятками видов. Интересно, что вместе с водорослями в таких местах живут и некоторые гетеротрофные грибы.

Рис.45. Подснежное развитие дубравных растений: слева — позеленевшие листья подснежника, пробившиеся сквозь снег; справа — ростки пролесника и чистяка, появившиеся из-под снега.
В период года, когда таяние снега невозможно, эти водоросли переходят в так называемое покоящееся состояние: прекращаются движение, рост и размножение, а активность других функций снижается до минимума.
Переход растения в состояние покоя может произойти вследствие непосредственного действия неблагоприятных условий. Но очень часто он совершается, как бы предупреждая приближающуюся смену жизненной обстановки. В этом случае наступление покоящегося состояния представляет выражение внутреннего, исторически выработавшегося и наследственно закрепленного ритма жизни растения. Растения, как часто говорят, готовятся к зиме.
Еще раньше времени наступления устойчивой отрицательной температуры замедляется, а потом и прекращается рост, растения сбрасывают листья, происходят некоторые физиолого-химические изменения, подготовляющие растение к зиме. Интересно, что в тропической зоне, в районах с периодически повторяющимися засухами, переход деревьев от состояния покоя к активной деятельности тоже происходит еще до начала периода дождей. Есть и у нас подобные явления. Выше упоминалось о подснежном росте некоторых травянистых растений (рис. 45), вспомним начало сокодвижения древесных пород, раннее цветение ольхи, орешника и другие ранневесенние явления.
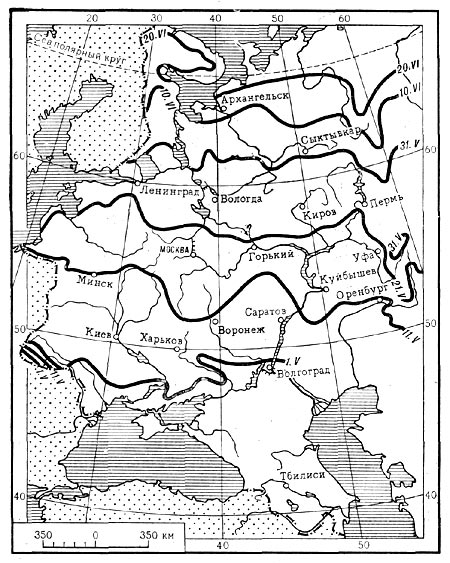
Карта 3. Сроки зацветания черёмухи в Европейской части СССР.
Тепло имеет большое значение для растений во время их развития в весенне-летний период. Многие широко распространенные растения служат предметом фенологических наблюдений. В разных пунктах на обширной территории ежегодно отмечают даты начала и конца цветения, распускания листьев и других фенологических фаз. Результаты многолетних наблюдений позволяют определить среднюю дату начала каждой фенологической фазы. Соединяя на карте плавными линиями пункты с одинаковыми сроками наступления каждой фазы, можно составить представление о том, где данная фаза наступает раньше, где позднее. На карте 3 видно, что в пределах Восточно-Европейской равнины чем севернее, тем позже зацветает черемуха. Объяснить это, очевидно, можно тем, что теплое время наступает тем позднее, чем севернее располагается местность. Аналогичные результаты установлены и для других фенофаз и различных растений.
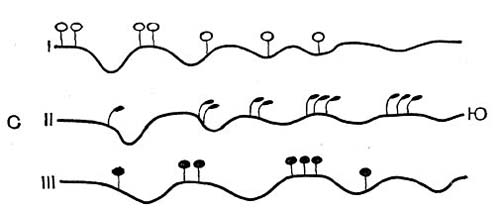
Рис.46. Правило предварения (схема): I -северный вид на юге переходит на северные склоны и в балки; II - южный вид на севере встречается лишь на южных склонах; III - вид с широким ареалом встречается на севере и на юге в экстразональных условиях.
Если по мере распространения растений в направлении полюса (или к вершинам гор) фаза цветения из-за похолодания климата в этом направлении будет наступать все позднее, то, принимая во внимание, что и продолжительность вегетационного периода в том же направлении укорачивается, можно прийти к выводу, что на некоторой географической широте (в горах — на некоторой высоте) расселяющееся растение будет располагать недостаточным временем для формирования плодов и созревания семян, благодаря чему дальнейшее расселение прекратится. В этом случае, очевидно, недостаток тепла сыграет роль фактора, ограничивающего географическое распространение растения. Тепловой фактор может сыграть аналогичную роль и по другим причинам. У многих деревьев в зимнее время происходит вымерзание ветвей — это обусловлено сочетанием низкой температуры с большой тратой воды на испарение. В результате древесные породы, испаряющие в зимнее время много воды, продвинулись в процессе расселения к северу меньше, чем древесные породы, испаряющие мало воды. Это, вероятно, надо считать одной из причин невозможности существования лесов в арктических широтах. То же произошло бы и с приземистыми растениями (кустарнички, травы), если бы их не защищал снег. Вообще же отрицательная температура вызывает отмирание отдельных органов и целых растений в результате механического разрушения тканей при образовании льда и вследствие иссушения, когда образовавшиеся в межклетниках кристаллы льда оттягивают из клеток воду. Но известно немало случаев, когда растения отмирают при снижении температуры, не достигающем 0° С. Это бывает с теми видами, становление которых совершалось в областях Земли, где не бывает отрицательной температуры. В этом случае происходит диско-ординация жизненных функций, что, в свою очередь, связано с нарушением равновесия работы ферментативных систем.
Оценивая влияние теплового фактора на гео-графизм растений, нельзя забывать о конкретных топографических условиях их жизни в данном географическом пункте. Дело в том, что даже в очень ограниченном географическом районе тепловые условия горизонтального водораздела и склонов разной экспозиции (особенно северной и южной) и разной крутизны бывают иной раз очень несходны. Часто случается, что растение, живущее на некоторой географической широте на горизонтальных плоских водоразделах (на «плакорах», в пла-корных или «зональных» условиях), встречается севернее только на более нагреваемых склонах южной экспозиции, а южнее данного района, наоборот, тяготеет к более прохладным, северным склонам (рис. 46). В двух последних случаях растение существует в экстразональных условиях. Не остаются без влияния на тепловой режим и субстрат его теплопроводность, теплоемкость, способность отражать тепло; ведь корни и вообще подземные органы в своей деятельности зависят от температурных условий не меньше, чем надземная часть растительного организма.
Таким образом, в пределах сколь угодно узкой климатической полосы в зависимости от разнообразия рельефа и тепловых свойств субстратов всегда наблюдается достаточно большое разнообразие местообитаний, отличающихся по тепловому режиму. Можно было бы ожидать, что в вечно теплых областях Земли тепловая обстановка не имеет существенного значения для растений. Однако это неверно. В тропической зоне, там, где наблюдается периодическая засуха, переход от засушливого времени к периоду дождей сопровождается незначительным повышением температуры, который вызывает переход растительности к буйной активности — как бы наступает весна. Даже очень небольшие температурные сдвиги в вечно теплых и одновременно вечно влажных областях, по-видимому, все же оказывают свое влияние на некоторые растения. Так, у одной орхидеи, живущей на Яве, наблюдается интересное явление: на большой территории цветочные почки, заложенные задолго до цветения, расцветают внезапно, в одно и то же утро, и цветение продолжается только один день. Причем оно наступает через девять дней, когда температура воздуха после регулярно повторяющейся грозы падает с 30 до 25° С.
Вода. В жизни растений вода играет весьма существенную роль. Она входит в состав живой плазмы растения. В виде водных растворов внутри растения передвигаются различные вещества, как воспринимаемые (тоже в растворенном состоянии) из окружающей среды, так и создаваемые растением в процессе его жизнедеятельности. Теряя воду при испарении, растение поддерживает восходящий водный ток и умеряет нагревание, защищаясь от перегрева. Но и находящаяся вне растения вода небезразлична для него. Количество атмосферных осадков и их распределение во времени, а также способность почвы (или другого субстрата) удерживать попавшую в нее воду в толще, доступной для корней, характеризуют условия водоснабжения растения. Облака в той или иной степени рассеивают, ослабляют свет, умеряют температуру и ее колебания. Туман рассеивает и поглощает свет и может быть также источником водоснабжения растений.
Влажность воздуха, определяемая содержанием в нем водяного пара, вместе с температурой определяют физическую обстановку потери воды при испарении. Даже в твердом состоянии вода небезразлична для растения. Лед, образующийся при промерзании почвы, практически перестает быть для растения источником воды. Снежный покров способствует сохранению тепла, образующегося в почве за счет дыхания почвенных организмов, и защищает прикрытые снегом живые части растения от неблагоприятного влияния низкой температуры воздуха и зимнего испарения, которое может привести к иссушению и отмиранию тканей.
Подавляющая масса растений, живущих на суше, в качестве основного источника воды использует почвенные и отчасти грунтовые ее запасы.
Источником воды в субстрате являются атмосферные осадки. Попадая в почву, вода под действием силы тяжести стремится проникнуть глубже, встречая, однако, на своем пути ряд препятствий. Таковыми могут быть прилипание к почвенным частицам (адсорбция), удержание воды структурными компонентами и коллоидными веществами почвы, поглощение живым населением почвы, наличие водонепроницаемых слоев. Таким образом, часть воды, проникшей в почву, задерживается в сравнительно ограниченном слое и в той или иной степени оказывается доступной растениям. Если водоупорный горизонт располагается на большой глубине, то скопляющаяся над ним грунтовая вода может быть доступна только растениям, способным развивать очень длинные вертикальные корни. Грунтовые воды передвигаются по уклонам водоупорных слоев и могут выходить на поверхность (ключи), становясь опять доступными растениям.
Однако не вся масса воды, попадая на поверхность почвы, проникает в нее. Если поверхностные слои почвы быстро насыщаются водой и перестают воспринимать (всасывать) ее, то избыток воды по склонам различных неровностей стекает в пониженные места и может попасть в постоянные потоки (реки), а по ним — в морские или не связанные с морем внутренние бассейны. Отсюда воде остается только один путь — в атмосферу, т. е. испарение.
Вода, проникшая в почву, испаряется с поверхности и, следовательно, также уходит в атмосферу. При этом если почва имеет хорошо развитую систему сообщающихся друг с другом капиллярных вместилищ, то на место испаряющейся воды поднимаются более глубоко находящиеся порции, которые также испаряются. Таким образом может происходить более или менее глубокое иссушение почвы. Следовательно, не вся вода атмосферных осадков, находящаяся в корнедоступном слое почвы, может быть использована растениями. Надо иметь в виду, что не обязательно вода атмосферных осадков достигает даже поверхности почвы. Орошая листья, вода дождей, роса, снег испаряются с поверхности крон и густого травяного покрова. Доля атмосферных осадков, уходящая в атмосферу, не достигнув поверхности почвы, может быть достаточно большой. Так, например, во вполне сомкнутых 80-летних ельниках под Москвой на кронах удерживается и уходит в атмосферу более 30% годовой суммы осадков. Немалая часть осадков испаряется в атмосферу и с поверхности густых травостоев. Количественные показатели в этом отношении очень изменчивы и зависят от густоты растительности, силы и продолжительности дождя. Непродолжительный и слабый дождь может и вовсе не проникнуть под полог леса.
Несмотря на то что не вся масса атмосферных осадков достигает почвы, все же большему количеству осадков соответствует большая вла-гообеспеченность территории в целом. С этой точки зрения для ботаника представляет большой интерес общая картина распределения осадков на Земле.
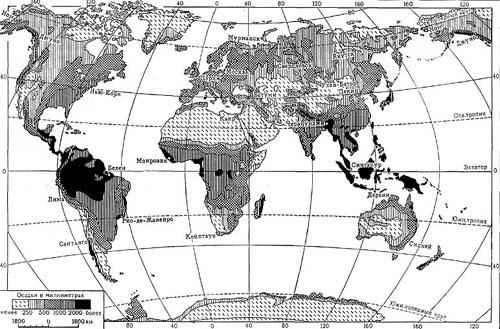
Карта 4.Распределение осадков на поверхности суши
Обеспеченность осадками зависит от географической широты. Сопоставление среднего за год количества осадков и географических широт показывает, что наибольшее количество осадков выпадает на экваторе; по обе стороны от него количество осадков снижается, достигая относительного минимума на широте 30° в южном и несколько севернее 20° в северном полушарии. Отсюда примерно до 50° в обоих полушариях намечается тенденция к повышению количества осадков, а далее по направлению к полюсу (в северном полушарии) их количество вновь убывает, снижаясь на 70° с. ш. всего до 21 см в год. Более точную и конкретную картину дает карта распределения осадков на Земле (карта 4). Ее интересно сопоставить с картой распределения главнейших типов растительности Земли (цветная карта 1 на вклейке). В северном полушарии обширны области пустынь и полупустынь, расположенные преимущественно в субтропических широтах, но простирающиеся в центральных частях материка и в области умеренных широт почти до 45° с. ш.; в южном полушарии пустынная и полупустынная растительность занимает место под тропиком в Африке и Австралии. Эти области получают осадков от 25 см и менее в год. В высоких широтах и в высокогорьях на карте растительности обозначены обширные территории почти с тем же количеством осадков, как в пустынях. Здесь нет древесной растительности — это главным образом тундры. Между этими двумя областями с малым количеством осадков, различающимися, однако, существенно по влажности воздуха, в северном полушарии, где находится основной массив суши, распространена обширная лесная полоса, покрытая хвойными и лиственными лесами.
В широкой полосе, ограниченной двумя тропиками и протянувшейся по меридиану более чем на 45°, растительность достаточно разнообразна. Здесь чередуются роскошные дождевые тропические леса, лишенные листвы в сухое время года, и обширные пространства саванн с господством в них злаков. Такое разнообразие растительности тоже связано с количеством осадков и особенно с их распределением. Дождевые тропические леса развиваются при 200 см и более осадков в год, но при условии отсутствия выраженного засушливого периода. Листопадные леса тропической зоны и саванны существуют при меньшем количестве осадков, но выпадающих неравномерно (в течение каждого года бывает один или два засушливых периода).
Эту чрезвычайно схематичную картину соответствия типа растительного покрова и распределения осадков можно детализировать. Необходимо показать, что изменчивость количества осадков, в пределах хотя бы лесной полосы Евразии, соответствует различию господствующих древесных пород, видовому богатству лесов, сложности их строения. Неодинаковы также и степи, как бы зажатые между полупустынями с одной стороны и лесами — с другой. Более детальный анализ покажет, что это разнообразие обусловливается (как и в тропической зоне) не только количеством осадков, но и их распределением в течение года, отсутствием или наличием засушливого периода и его продолжительностью. Словом, распределение растительности не только на Земле в целом, но и в пределах каждой зоны и каждого из материков приводит к бесспорному выводу, что влажность климата представляет одно из важнейших условий растительной жизни, она определяет общий характер растительности.
Это заключение получает подтверждение и в еще более тонких деталях распределения растений. Рассматривая распределение растений и их сочетаний в пределах однородного в отношении влажности климатического района, нетрудно убедиться в том, что неодинаковая водообеспеченность растений играет нередко ведущую роль в их размещении по территории. Так, например, наблюдая где-нибудь в средней части лесной полосы в пределах Восточно-Европейской равнины, в местности с более или менее развитым рельефом, распределение еловых лесов по склону к небольшому водотоку, можно заметить, что наиболее высокие и вместе с тем наиболее сухие участки заняты ельниками с брусничным покровом. Ниже их располагаются ельники с травянистым покровом из кислицы, майника и ряда других трав, а вблизи водотока, в наиболее влажных местах, найдем так называемый прирученный ельник с достаточно обильным сочным травяным покровом из папоротников, высоких трав, имеющих обычно крупные и широкие листья. Наблюдения за растениями, сопровождающими в названных типах елового леса ель (поскольку они встречаются и вне ельников), показывают, что травянистые растения приручейных ельников практически всегда связаны с более увлажняемыми местами, чем те, на которых в массовых количествах растет брусника, а кислица предпочитает места более обеспеченные водой, чем брусника. Отсюда можно сделать вывод, что ряд ельников, закономерно сменяющих друг друга по склону, в большой степени соответствует возрастанию увлажнения.
Факты зависимости распределения целых комплексов растений на разных уровнях склонов многочисленны. Подобную картину можно наблюдать не только в лесах, но и в степях, на суходольных и заливных лугах, даже на болотах. Анализ конкретных высотных экологических рядов (так называют природные последовательности закономерных сочетаний растительных сообществ, соответствующих изменению какого-нибудь фактора в убывающем или нарастающем порядке) требует некоторой осторожности, так как не всегда высотный ряд обусловлен нарастанием или убыванием водообеспеченности. Бывает, что распределение растительности по высоте склонов определяется иными почвенно-грунтовыми условиями, например снабжением почвы кислородом, распределением минеральных солей и пр.
Если целые комплексы растений размещаются соответственно условиям водоснабжения и в масштабе всей суши, и в пределах более ограниченных климатически однородных территорий, то, очевидно, и отдельно взятые виды, и составляющие их особи также живо реагируют на этот фактор. Свойства растения, в частности его отношение к водообеспеченности, складывались в процессе эволюционного становления и у каждого вида соответствуют более или менее узкой амплитуде изменчивости среды. В отличие от животных подавляющая масса растений лишена способности активно изменять свое положение в пространстве. Растение не имеет возможности «отыскивать» лучшее для себя место, что доступно многим животным. Следовательно, каждое растение может жить в данном месте, если существующие здесь условия достаточны для него.
Современная флора суши состоит в основном из цветковых растений, строение тела которых наиболее сложно и в высокой степени соответствует возможности существования в двухфазной среде (твердый субстрат и воздушное окружение). Эта приспособленность к жизни на суше — результат длительного эволюционного процесса. Мысленное восстановление эволюционного пути (на основе сохранившихся памятников), пройденного организмами до той степени совершенства, которого достигли цветковые растения, приводит к выводу, что их предками были водоросли.
Водоросли обычно живут в воде, вся поверхность их тела соприкасается с водой, и таким образом они не сталкиваются с проблемой водо-обеспечения. Но если водоросли стали предками наземных растений, то это значит, что некоторые из них должны были некогда «выйти на сушу», преодолеть отсутствие водного окружения и связанную с ним опасность высыхания. Водные предки наземных растений не испаряли воду. Выход из воды, приведший к непосредственному соприкосновению с воздухом, был едва ли не самым трудным шагом в процессе формирования наземных растений. Естественно возникает вопрос: могли ли водорослевые предки преодолеть барьер между обстановкой полной обеспеченности водой и условиями, когда организм на протяжении своей жизни, по крайней мере периодически или эпизодически, оказывался вне водного окружения? Ответ на этот вопрос положительный. В современный период в приливно-отливной полосе моря водоросли регулярно остаются вне воды на протяжении некоторой части суток. Если водоудерживаю-щая способность протоплазмы их клеток достаточно велика, они не умирают и, говоря образно, набирают силы в часы прилива. Один шаг отделяет поселенцев этой, как ее называют, литоральной полосы от супралиторальной, находящейся выше наиболее высокого прилива, где имеется, однако, регулярно повторяющееся орошение брызгами волн прибоя. Естественно, в этих, еще более жестких условиях водоснабжения древние иммигранты суши должны были приобрести и более высокие водоудерживаю-щие свойства плазмы и способность ее оставаться живой достаточно длительное время при отсутствии орошения.
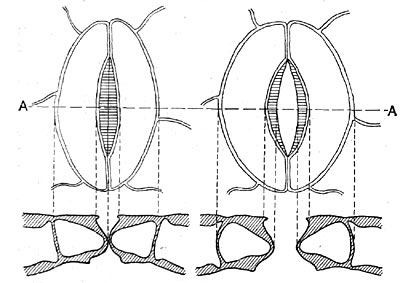
Рис. 47. Устьица: сверху - вид с поверхности; внизу - в разрезе; слева - в закрытом состоянии; срава - в закрытом состоянии. (А-А - линия разреза)
Все эти растительные организмы, более или менее хорошо переносящие перерыв в водоснабжении, имели сходство в том, что содержание в них воды — их оводненность — почти целиком зависела от окружающей среды. Недостатку воды растительные организмы могли противопоставить способность пережить период безводья, переход в крайнем положении в состояние замедленной скрытой жизни, как бы в оцепеневшее состояние: Они еще бессильны были сами поддерживать свою оводненность на достаточно высоком уровне и в течение длительного времени. Их оводненность слагалась во времени как чередование то большего, то меньшего содержания воды. Картина оводнен-ности была изменчивая, зависящая от состояния ер-еды, как бы пестрая. Это дало основание назвать растения с подобным ритмом оводнен-ности пойкилогидридными (греч. «пойкилос» — пестрый, «гидро» — вода).

Таблица 15. Жизненные формы (водные травы): вверху (слева направо) — желтая кубышка, белая кувшинка, калла (белокрыльник); внизу — виктория регия.
Интересно, что и в современную эпоху достаточно широко распространены растения этой категории. На почве и внутри ее живет немало аэрофильных водорослей — пойкилогидрпдов, тело которых непосредственно соприкасается с воздухом. Некоторые из них живут на коре деревьев, камнях, скалах и т. д. Но, что самое интересное, есть водоросли (сине-зеленые), населяющие поверхность почвы в полупустынях и даже в пустынях. Они образуют слизистую массу, в которую погружено нитчатое тело водоросли и которая удерживает в себе воду. Водоросль вегетирует, пока слизистая масса не высохнет, превратившись в хрупкую корочку, и пребывает в таком состоянии, пока вновь не набухнет от дождей или от талой воды весной. Водоросли, живущие ныне вне воды, не являются, вероятно, прямыми потомками тех первичных пойкилогидридов, которые существовали в эпоху формирования наземной растительности. В ряде случаев можно предполагать родственные связи современных вневодных (аэрофильных) водорослей с современными же, но живущими в воде. В таком случае в современную (геологическую) эпоху природа как бы повторяет опыт осуществленного впервые в незапамятные времена первичного заселения суши растениями, чем и подтверждается ранее описанная схема иммиграции водорослей на сушу.
Пойкилогидридность известна не только у водорослей. Она свойственна бактериям, миксомицетам, грибам, лишайникам, т. е. низшим слоевцовым растениям; она распространена также у многих мхов, у заростков папоротников, а иногда даже бывает и у листьев папоротников. Более того, есть, по-видимому, и некоторые цветковые пойкилогидриды. Но не на основе пойкилогидридности формировалось основное разнообразие современного растительного мира суши. Главную массу растений современной флоры составляют не пойкилогидриды, агомоиогидриды (от греч. «гомойос» — одинаковый), т. е. растения, способные сами поддерживать свою оводненность в пределах некоторого уровня и в известной мере (в этом отношении) независимые от внешней среды.
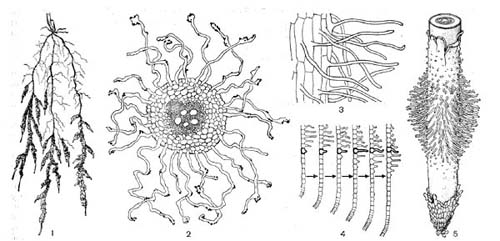
Рис.48. Корневые волоски: 1 — четырехнедельного проростка пшеницы с прилипшими к -ним частицами почвы; 2 — поперечный разрез корня, видны корневые волоски с прилипшими к ним частицами почвы; 3 — клетки ризодермы, образовавшие корневые волоски; 4 — формирование корневых волосков и перемещение их вслед за ростом корня; 5 — кончик корня с'корневыми волосками и чехликом.
Относительная устойчивость оводненности представляет результат балансирования траты воды на испарение и добывания ее в количествах, обеспечивающих потребность организма в воде. Первое определяется выработкой покровных тканей, среди которых особо важное значение имеет кожица, или эпидермис. Он покрыт слоем затрудняющей испарение кутикулы, и в его составе имеются саморегулирующие межклеточные отверстия — устьичные щели (рис. 47). Работа устьиц, т. е. расширение их щелей и смыкание замыкающих клеток, определяется рядом обстоятельств, среди которых одно из ведущих — сама степень оводненности растения: при падении оводненности устьица закрываются, при повышении — щель их вновь расширяется.
Добывание воды у гомойогидридных растений обеспечивается в основном корнями, вблизи растущих окончаний которых образуется множество корневых волосков, обладающих в совокупности огромной наружной поверхностью. Через нее и поступает в растение вода, имеющаяся в почве (рис. 48).
Связь между аппаратом, добывающим воду и расходующим ее, сложная. Недостаток воды вызывает повышение всасывающей деятельности корней и в то же время приводит к закрыванию устьиц. Но деятельность устьиц осложняется тем, что раскрытие устьичной щели регулируется также освещением — результат того, что через устьица не только выделяется водяной пар, но и осуществляется поступление в растение углекислого газа и выделение кислорода (фотосинтез).
В общих чертах можно сказать, что у го-мойогидридов по мере потери воды клетками возрастает энергия, с которой они всасывают воду, т. е. растет их сосущая сила. При достижении высокой оводненности сосущая сила снижается.
Благодаря этому гомойогидридные растения способны поддерживать в некоторых пределах свою оводненность.
Будучи основными растениями современной суши Земли, гомойогидриды населяют территории и с разным климатом (количество осадков, условия испарения), и в пределах однородного климата — места то сильно увлажненные, то очень сухие. Естественно, растения, живущие в сухих местах, могут отличаться от растений, населяющих влажные места, рядом своеобразных приспособительных особенностей, как внутренних, связанных со свойствами протоплазмы, так и структурных, усиливающих возможность контроля расходования воды и водо-поглощения. Такие отличия действительно существуют, а структурные особенности часто настолько выражены, что по внешнему облику растения нередко можно определить, связано ли оно с сухими, влажными или с промежуточными условиями.
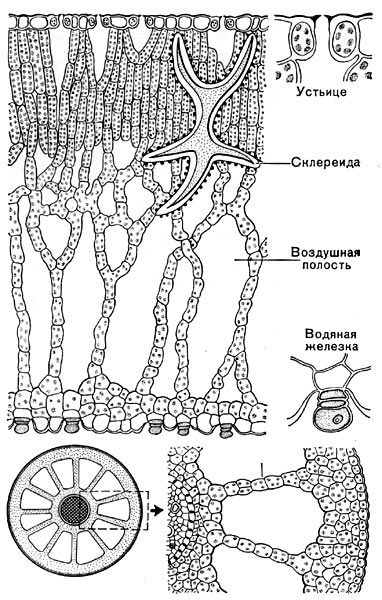
Рис.49. Анатомическое строение некоторых водных растений. Вверху — поперечный разрез через плавающий лист желтой кувшинки; видны обширные воздушные полости; отдельно изображены устьице и водяная железка. Внизу — поперечный разрез стебля урути, справа — при большом увеличении.
С точки зрения отношения растений к обеспечению водой выделяются следующие экологические группы: ксерофиты, мезофиты, гигрофиты, гидрофиты, гидатофиты.
Гидатофитами называют водные растения, целиком или почти целиком, т. е. большей частью тела, погруженные в воду. Примером может служить водяная чума (элодея), рдесты, кувшинки и др. Гидрофиты — водонадземные растения, прикрепленные к почве. Они начинают свое индивидуальное развитие и ежегодное возобновление побегов будучи погруженными в воду, но во взрослом состоянии верхние части побегов выступают над поверхностью воды. Гидрофиты обитают в прибрежьях водоемов, на неглубоких местах, но могут жить и на обильно увлажненной почве вдали от водоема. В отличие от гидатофитов гидрофиты имеют ясно выраженные механические ткани и водопроводящую систему. Обычно у них хорошо развита система межклетников и воздушных полостей, по которым воздух, поступающий через устьица, проникает и в нижние части растения, скрытые в перенасыщенном водой субстрате. Для гидатофитов и гидрофитов не обязательна влажность климата, так как и в областях, недостаточно обеспеченных осадками, даже в пустынях, могут существовать водоемы, полностью удовлетворяющие потребность таких растений в воде.
Гигрофиты — растения, живущие на обильно увлажненных почвах и, что особенно характерно, при высокой влажности атмосферы. Последнее обстоятельство играет в жизни гигрофитов важную роль, так как благодаря этому замедляется, а то и вовсе исключается испарение, что отзывается на восходящем (от корней к листьям) токе воды в растении, приводит к замедлению снабжения переносимыми водой внутри тела растения минеральными питательными веществами. Развитие обширных внутренних полостей у гигрофитов должно способствовать усилению потери воды клетками — вода выделяется в виде пара в межклетные полости, а из них через устьица наружу. У растений этой категории кутикула тонкая и ее роль в ограничении внеустьичного испарения сводится к минимуму. Листовые пластинки часто бывают тонкими, из незначительного числа слоев клеток, благодаря чему большая часть клеток листа непосредственно соприкасается с воздухом, что, естественно, способствует водоотдаче. Иногда, как, например, у некоторых папоротников, живущих в тени влажных тропических лесов, листовая пластинка может состоять всего из двух, даже из одного слоя клеток. Тем не менее и эти приспособительные особенности не всегда достаточны для поддержания тока воды. Для гигрофитов очень характерно наличие на листьях особых железок — это так называемые гидатоды или водяные устьица, через которые происходит активное выделение воды в капельно-жид-ком состоянии, что позволяет поддерживать движение воды в растении и поглощать новые ее порции из почвы даже в случае, если воздух насыщен водяным паром. Из числа гигрофитов нашей флоры можно назвать чаровницу, или цирцею, адоксу, недотрогу (рис. 49), болотный подмаренник.

Таблица 16. Жизненные формы (травы наземные — паразитные и насекомоядные): вверху (слева направо) — петров крест, непентес, росянка; внизу — раффлезия. (лесные травы и кустарники, мхи и стелющиеся цветковые).
Мезофиты — растения, обитающие при достаточном (не избыточном и не слишком ограниченном) увлажнении. Типичные мезофиты обычно связаны и с умеренными тепловыми условиями, и с хорошими условиями минерального питания. Одни мезофиты — лесные растения, другие — луговые, живущие вообще на незатененных местах, что в известной мере отзывается на их облике. Среди мезофитов нередки древесные растения и кустарники, такие, как липа, береза, лещина (орешник), крушина и многие другие. Но еще больше среди них трав: клевер красный, тимофеевка, костер, луговая овсяница, нивяник и многие другие луговые растения, ландыш, копытень, сныть и целый ряд других лесных растений. В целом категория мезофитов очень обширна. Резкую границу между ними и гигрофитами установить трудно, поэтому приходится выделять переходную категорию растений — гигромезофиты. Невозможно также четко разделить мезофиты и ксерофиты — растения засушливых мест. Здесь также отличают ксеромезофиты (более близкие к мезофитам) и мезоксерофиты (больше приближающиеся к ксерофитам).
Ксерофиты едва ли не наиболее разнообразная и трудноопределимая экологическая группа растений. Эти растения нельзя характеризовать географически, как растения засушливых областей, так как в любой засушливой области могут быть местные условия, обеспечивающие существование растений иного типа, а иногда, наоборот, в областях с достаточным количеством осадков создаются местные условия недостаточного водообеспечения, доступные только ксерофитам. Поэтому лучше считать ксерофитами растения, живущие на местах, сухих настолько, что добывание и экономия воды требуют особых к этому приспособлений. Приспособления этого рода можно объединить в три категории: ограничивающие испарение; усиливающие добывание воды при ее недостатке в почве; позволяющие создавать запасы воды на время длительного перерыва водоснабжения.
Сокращение испарения достигается разными способами. Растение образует мелкие (сравнительно с близкими видами нексерофитами) листья или листья вовсе редуцированы. Мощная кутикула, покрывающая толстые наружные стенки клеток эпидермиса, практически полностью исключает внеустьичное испарение (рис. 50). Уменьшению испарения способствует также развитие воскового налета. Кроме того, блестящая поверхность кутикулы отражает солнечные лучи. Этот же эффект достигается при помощи густого опушения из бесцветных волосков, образованных пустыми безводными клетками, рано теряющими свое содержимое. Опушение играет и другую роль: между волосками создается зона затишья, и в ней задерживается водяной пар, что замедляет испарение. Аналогичный эффект достигается положением устьиц в углублениях, где также задерживается выделенный устьицами пар. Наблюдаемое у ряда ксерофитов свертывание листьев на сторону, с которой располагаются устьица, также приводит к обогащению воздуха паром внутри образовавшейся трубочки. Наконец, плотное сложение паренхимной ткани листьев, т. е. малое количество межклетников, также умеряет потерю воды. Все эти структурные особенности часто сопровождаются обильным содержанием в листе и других органах ксерофита опорных (механических) тканей, мешающих увяданию.
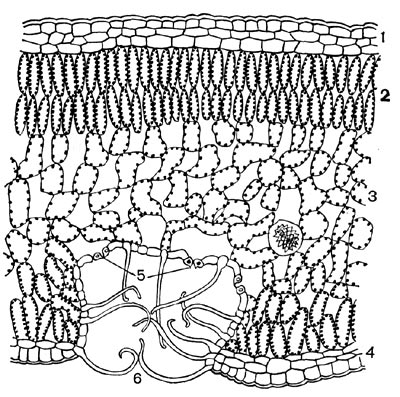
Рис.50. Разрез листа олеандра. Устьица в углублениях на нижней стороне листа, закрытые волосками: 1 — верхний эпидермис; 2 — палисадная паренхима; 3 — губчатая паренхима; 4, — нижний эпидермис; 5 — устьица; в — волоски.
Повышенная способность ксерофитов добывать воду связана в основном с корнями. В целом у ксерофитов бывает более мощная, чем у мезофитов и гигрофитов, корневая система. Можно различить по крайней мере три типа ее организации.
У некоторых ксерофитов особенно энергично растет главный корень. Направляясь вертикально вниз, он достигает у взрослого растения тех глубоких горизонтов, где субстрат постоянно влажен, не просыхает. Такой корень может достигнуть и грунтовых вод или по крайней мере дорасти до зоны их капиллярного поднятия. Установлено, что иногда главный корень проходит сквозь толщу, измеряемую метрами и даже десятками метров. Так, у желтой люцерны главный корень достигает 6—8 м глубины, а у живущей в полупустынных и пустынных районах верблюжьей колючки стержневой корень достигает 18—20 м глубины (рис. 51). В достаточно влажных слоях стержневой корень обильно ветвится, и благодаря этому растение обеспечивает себя водой.
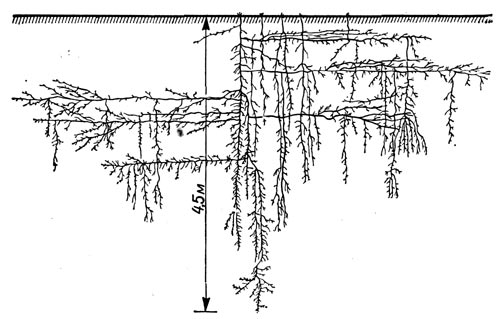
Рис.51. Корневая система верблюжьей колючки в возрасте 17 месяцев, достигшая глубины 4,5 м.
Второй тип организации корневой системы характеризуется наличием множества корней (обычно придаточных), которые, образуя раз-ветвления нескольких порядков, густо пронизывают весь объем почвы, охватываемый корневой системой в целом. Такое строение корневой системы, характерное, например, для дерновинных злаков (рис. 52), дает возможность растению очень интенсивно использовать почву как источник воды (а также и элементов минерального питания). Такие корневые системы наблюдаются у мезоксерофильных и даже у мезофильных растений. Они свойственны, например, ржи, пшенице, овсу и ряду других хлебных растений и некоторым сорнякам. Изучение корневой системы ржи показало, что совокупность длины всех корней одного экземпляра в посеве, при расстоянии между особями 15 см, составляет почти 1 км. Если растение живет вне посева, то, не встречая препятствий со стороны соседей, совокупная длина корней достигает почти 80 км. Еще большей мощности может достигнуть корневая система у овсюга (более 86 км).
Все эти цифры возрастают во много раз, если к ним прибавить совокупную длину корневых волосков.
Третий тип организации корневой системы — поверхностная система. Она также слагается из множества разветвлений, главным образом из придаточных корней, которые, однако, не углубляются в почву, а располагаются на небольшой глубине. Такая корневая система может активно обеспечивать наземные части растения водой только во время выпадения осадков, которые улавливаются сетью корней, всасываются ими, как только вода попадает в почву. Такого рода корневые системы эффективны в том случае, если растение способно запасать воду, сохраняя ее в своем теле и медленно расходуя во время длительной засухи. Каждый из трех типов корневых систем ксерофитов ориентирован на разные источники воды. Растения с глубоко проникающим стержневым корнем используют глубинную воду, представляющую хоть и труднодостижимый, но достаточно надежный постоянный источник водоснабжения. Растения с поверхностным типом корневой системы ориентированы на максимальное использование осадков почти в момент их выпадения. Такие растения имеют возможность в большей степени использовать осадки, чем растения с глубинной корневой системой. Мощная корневая система, интенсивно пронизывающая весь доступный растению объем почвы, занимает промежуточное положение. Она использует воду, проникшую в почву и задержанную ею, т. е. не ушедшую в глубину, где залегают грунтовые воды.
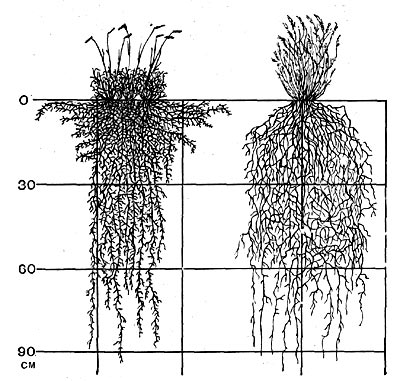
Рис.52. Корневая система Aristida purpurea (справа)и Bouteloua gracilis (слова).
Эти три варианта корневых систем не исчерпывают их разнообразия, тем более что часто у одного и того же растения можно встретить комбинации разных принципов строения. Так, например, у дерновинных злаков наших степей корневая система построена по второму типу, но весной и после выпадения дождей у них развивается масса поверхностных корней, которые значительно повышают эффективность восприятия воды.
Все эти особенности гомойогидридных ксерофитов характеризуют главнейшие морфологические особенности ксерофитов. Но им присуще и известное физиологическое своеобразие, ведущей чертой которого, свойственной большинству ксерофитов, является повышенное осмотическое давление. С ним тесно связаны пониженная оводненность и повышенная сосущая сила. Всякое изменение в водном балансе растения отражается на его оводненности, а это приводит к изменению осмотического давления и зависимой от него сосущей силы.
Рассматривая жизнь растений при недостатке осадков и низкой относительной влажности воздуха, следует учитывать, что водный режим растений тесно связан с другими функциями, в частности с фотосинтезом. Так, сокращение потери воды благодаря закрытию устьиц влечет за собой сокращение поступления углекислого газа и, следовательно, снижение фотосинтеза, приводящее к ограничению или даже прекращению роста.
Растения, способные жить в засушливых условиях, по-разному преодолевают их. У некоторых практически нет структурных приспособлений для перенесения недостатка влаги. Их существование в засушливом климате возможно вследствие того, что они, находясь в течение всей засухи в состоянии покоя, быстро переходят к активной жизнедеятельности и в течение короткого промежутка времени (11/2 — 2 месяца) завершают годичный цикл развития. Такие растения называют эфемерами (однолетние, сохраняющиеся в сухое время только в виде семян) и эфемероидами (многолетние, сохраняющиеся в виде луковиц, клубней, корневищ, прекращающих рост на сухое время и переживающие его на некоторой глубине под поверхностью почвы). Эфемерные растения построены, как мезофиты. Это — мезофиты, «убегающие» от засухи.
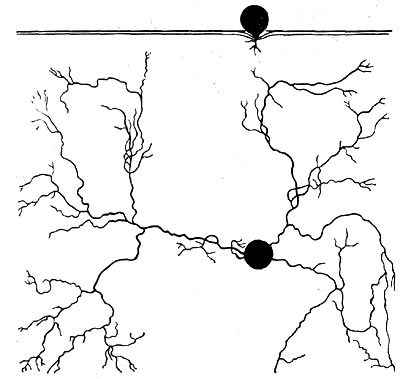
Другую своеобразную категорию растений, переносящих засушливые условия, представляют суккуленты. Суккуленты способны накапливать в своих тканях много воды и медленно расходовать ее. Запасаемая вода сохраняется в листьях таких растений, как агавы, алоэ, очитки и др., которые называются листовыми суккулентами. Стеблевые суккуленты содержат воду в стеблевых органах (кактусы, кактусовидные молочаи, стапелии и др.); причем листья у таких растений редуцированы, превращены в колючки; кутикула очень толстая, покрытая воском; устьица немногочисленные, погруженные и в период засухи закрытые, вследствие чего рост у этих растений очень медленный. Осмотическое давление низкое. Количество воды, скопляющейся в теле, может быть очень велико. Например, некоторые кактусы североамериканских пустынь накопляет от 1000 до 3000 л воды. Довольно широко распространены суккуленты, запасающие воду в подземных органах: в клубнях, корневищах и даже в корнях.
Прочие ксерофиты можно разделить на две недостаточно резко разграниченные группы — эуксерофиты и гемиксерофиты. Иногда первую группу, в свою очередь, подразделяют на две ступени — эуксерофиты и ксерофиты. Среди же гемиксерофитов часто различают мезоксерофиты и ксерофиты.
Эуксерофиты — ксерофиты в собственном смысле слова, характеризуются пониженной транспирацией, особенно в часы наибольшей сухости воздуха. Гемиксерофиты (полуксерофиты) — растения, способные существовать в засушливых условиях благодаря достаточно мощной корневой системе. Эти растения испаряют много воды, так как в единице их поверхности содержится большое количество устьиц и сильно развита система жилок в листьях. У некоторых гемиксерофитов сумма длин всех жилок в пределах 1 см2 может превышать 1200 мм. Число устьиц на нижней поверхности листа у каждого из этих растений приближается к 450 на 1 мм2. Гемиксерофиты могут довольно энергично расти и сохранять свежую листву даже в наиболее сухое время.
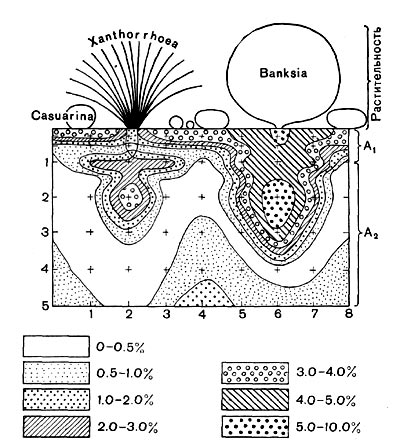
Рис.54. Распределение влажности в почве маккии Южной Австралии на 24/1II после небольшого дождя. Вертикальный и горизонтальный масштаб одинаковы: каждое деление соответствует 1 англ, футу (около 30 см). Объяснение в тексте. A1 и А2 — почвенные горизонты.
В случае недостатка воды такие растения иногда способны уменьшать испаряющую поверхность, сбрасывая листья или заменяя их более мелкими. Гемиксерофиты могут тратить воду на испарение в большем количестве, чем мезофиты.
Таким образом, в пределах групп растений, отличающихся отношением к водообеспеченно-сти, имеется гамма переходов от совершенно неиспаряющих погруженных гидатофитов к полуназемным и малоиспаряющим наземным растениям (гигрофитам), далее к более испаряющим — мезофитам — и наконец опять к малоиспаряющим эуксерофитам. Другими словами, максимально и минимально обеспеченные водой растения испаряют ее меньше, чем растения, занимающие промежуточное положение.
Мы рассмотрели основные черты зависимости растений от влагообеспеченности, которая в конечном счете создается сочетанием абиотических факторов: количеством осадков, их распределением, свойствами субстрата, температурными условиями. Возникает вопрос: влияют ли сами растения на водные условия их жизни? Такое влияние, несомненно, есть. Во всяком случае, нельзя, например, отрицать, что, по мере того как запас влаги, образовавшийся в почве, используется растениями, происходит некоторое иссушение субстрата. Оно может быть ликвидировано, если пройдет дождь, и временное иссушение субстрата может пройти бесследно для растений. Если количество осадков невелико и они выпадают главным образом в начале вегетации, то к середине летнего сезона при достаточно высокой температуре в почве неминуемо должен возникнуть существенный недостаток воды. Действительная картина влияния растений на влажность почвы, вероятно, довольно сложная. На рисунках 54 и 55 показан результат исследования содержания воды в песчаной почве, занятой жестколистными кустарниками в Южной Австралии. На рисунке 54 изображено распределение влажности после выпадения дождя (24 мм) 24 марта (в середине засушливого периода) в почве, которая до того была равномерно сухой. Данные рисунка 54 позволяют заключить, что по крайней мере два растения — ксанторрея и банксия — очень сильно повлияли на распределение этой порции влаги: под ними, особенно под банксией, почва оказалась наиболее увлажненной. Влияние тех же растений на распределение влаги после окончания влажного периода, в начале летней засухи, показано на рисунке 55. Под банксией на глубине от 60 см и более создалось пятно сильного иссушения (влажность 0,5—1,0%), а по мере приближения к поверхности влажность возрастает и под самым кустом близ корневой шейки достигает 4,0—5,0%, т. е. того уровня, на котором влажность держится в почве на глубине 180—190 см от поверхности. При объяснении отраженной в этих рисунках влажности почвы необходимо связывать ее с деятельностью указанных растений.
Благодаря испарению воды непосредственно с поверхности растительного покрова почва под густой растительностью получает меньшее количество атмосферной воды, чем то, которое улавливается дождемерами. Растительность живет как бы в климате с иным количеством осадков. В некоторых случаях это может иметь, по-видимому, существенное значение.
Ранее было указано, что, в частности, еловый лес «не допускает» до почвы примерно 30% всего количества осадков. Этим предупреждается переувлажнение почвы, которое может привести к заболачиванию; ель же не переносит заболачивания. В глубине леса, так же как у поверхности земли, под достаточно густым травяным покровом создается повышенная влажность воздуха, которая влияет на проростки и ювенильные особи деревьев (в лесу) и трав, образующих сомкнутую массу на лугах, а также на растения, во взрослом состоянии низкорослые и занимающие место вблизи почвы
Примеров воздействия отдельных растений, особенно целых их сообществ, на среду можно было бы привести много. Все они свидетельствуют о том, что растительные организмы в совокупности создают свой особый, внутренний климат — микроклимат. Их деятельность отражается на световом режиме, температурных условиях, влажности воздуха, распределении влаги в почве. Можно к этому прибавить, что под влиянием растительности изменяются ветровые потоки. Например, над кронами дубового леса в течение почти 500-часовых наблюдений только в 10% случаев был констатирован штиль (скорость ветра менее 0,7 м/сек), в то время как непосредственно над почвой этого леса штилевые промежутки времени составляли 98%.
Нередко думают, что лес как бы притягивает осадки, а после вырубки лесов осадков становится меньше. Из этого делают вывод, что характер растительности имеет прямое влияние на формирование местного климата. Действительно, некоторые данные, подтверждающие такое положение, имеются. Только увеличение количества осадков в облесенных местностях по сравнению с необлесенными невелико и достигает не более 6%. Есть также данные, что влажность воздуха в ближайших окрестностях лесных массивов бывает повышенной. Более значительное участие леса в формировании влажности климата заключается в следующем. Массы воздуха, движущегося от Атлантического океана к востоку, проходя над Гольфстримом, обогащаются влагой. Двигаясь над материком, воздух теряет влагу в виде осадков, но он может вновь обогатиться водяным паром за счет испарения с поверхности Земли. Наиболее мощным испарителем является лесная растительность, которая и становится поставщиком атмосферных осадков для районов, располагающихся восточнее и юго-восточнее по пути движения океанического воздуха, пришедшего с запада. Испаряя воду, растительный покров может вернуть в атмосферу тем большую часть полученной из нее воды, чем мощнее аппарат, испаряющий воду, и чем меньше атмосферных осадков уходит в виде стока за пределы испаряющего массива.
Лесная растительность не только является наиболее мощным испарителем влаги, но и в наибольшей степени предохраняет воду Атмосферных осадков от поверхностного стока. Состав и движение воздуха. Газовая фаза среды обитания наземных растений служит для них резервуаром углекислого газа. Его содержится в воздухе в среднем, всего 0,03%, но его потребление в процессе фотосинтеза постоянно пополняется за счет дыхания животных, микро- и макроорганизмов почвы и животного населения океана, сжигания огромных масс органического вещества (топливо) и, наконец, выделений из недр Земли. В результате в атмосфере поддерживается такое количество углекислого газа, которое сравнительно мало (в абсолютных показателях) отклоняется от средней нормы. Более или менее заметные отклонения ограничены и в пространстве, и во времени.
Содержащийся в воздухе кислород (21%) используется растениями и животными при дыхании.
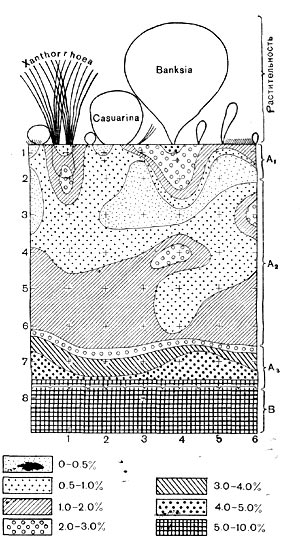
Рис.55. Распределение влажности в почве в том же месте (см. рис. 54) после периода дождей, к началу засушливого времени, 13/XII. Объяснение в тексте.
Наиболее обильный (79,5%) газ атмосферы -азот — подавляющей массой растений не усваивается. Нуждаясь в этом химическом элементе, растения потребляют его в виде азотсодержащих соединений. Только очень немногие растения (некоторые бактерии, сине-зеленые водоросли) удовлетворяют свою потребность в азоте за счет запасов этого газа в воздухе. Значение таких азотфиксирующих растений огромно, они обогащают почву азотсодержащими соединениями.
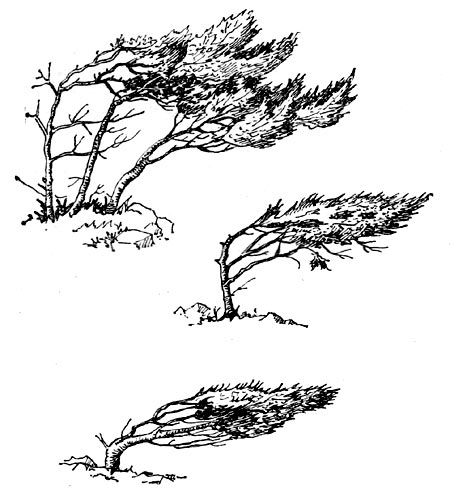
Рис.56. Влияние ветра на обычно правильную крону можжевельника Juniperus excelsa. Крым, берег моря; флагообразная крона.
Иногда существенную роль играют ядовитые газовые примеси, имеющие значение главным образом в местностях с высокоразвитой индустрией. Наиболее существенная роль принадлежит двуокиси серы. Это кислородное соединение серы сильно подавляет рост растений. Ядовитое действие начинается уже при содержании одной объемной части двуокиси серы в 200 000—500 000 объемов воздуха. Во многих промышленных центрах наблюдается значительно более высокая концентрация этого газа.
На жизнь растений большое влияние оказывает и движение воздуха. Роль его разнообразна. Общеизвестно, что воздух является важным фактором распространения растений, переносчиком семян, плодов и тем более спор растений и играет большую роль в опылении растений все голосеменные растения ветроопыляемы, ветром же переносится пыльца и многих покрытосеменных растений. Кроме того, ветер действует на растения иссушающе, вследствие того что во время ветра порции воздуха, обогащенные паром, выделенным растением, уносятся и заменяются более сухими. Но новые порции воздуха могут принести и новые дозы углекислого газа. Ветер несколько охлаждает растение, однако если воздух содержит много сильно нагретых пылевых частиц, то температура вокруг растения может значительно повыситься. В целом влияние ветра сложно и разнообразно. Ветер иногда вызывает значительные изменения в облике растения. Это касается главным образом одиночных деревьев, растущих на берегу моря и в горах, где преобладают ветры одного направления и большой силы; иссушающее воздействие ветра сказывается в основном на наветренной стороне кроны. Здесь иссушение листьев может значительно возрастать вследствие изгибания листьев очень сильным ветром. При этом воздух, заполняющий межклетники, выжимается наружу, благодаря чему иссушение резко возрастает. Сильное иссушение приводит к замедлению роста ветвей с наветренной стороны дерева, и в итоге формируется односторонняя, асимметричная крона, иногда вовсе лишенная ветвей с наветренной стороны (рис. 56). Ураганные ветры не только обламывают крупные ветви, но и валят даже целые деревья. Ураган может погубить лес на большой площади лесопокрытой местности.
Механическое и иссушающее действие постоянных ветров в процессе эволюции привело к возникновению своеобразных жизненных форм растений.
Следовательно, чтобы оценить конкретную климатическую обстановку в каком-нибудь географическом пункте, надо мысленно обобщить действие и взаимодействие всех климатических факторов и суммировать все изменения погоды в данном месте в течение года. Приближенно это можно определить с помощью климадиа-грамм. При построении климадиаграмм учитывают наиболее мощные и важные для растений климатические характеристики — влажность и тепловые условия. Так, изображенные на рисунке климадиаграммы для Москвы, Каира и Дуалы (Камерун) показывают резкое своеобразие климата каждого из этих пунктов. Конечно, климадиаграммы не учитывают еще ряда экологически важных особенностей климата: общее количество солнечной энергии, число безоблачных дней в году и их распределение, господствующие ветры, их направление по сезонам года и скорости и т. п. Но все эти показатели могут быть использованы дополнительно для выявления более тонких отличий между климатами со сходными климадиаграммами.
Эдафические факторы определяются свойствами субстрата. Субстрат может быть незначительно затронут процессами физического выветривания или в результате воздействия организмов поверхностный слой невыветренной породы ее превращается в почву. Почва — тело сложного состава с закономерным взаимоположением отличающихся друг от друга слоев (или горизонтов). Термин «почва» применяется иногда не только к почвам сформированным, но и формирующимся и даже к субстратам, еще не затронутым почвообразовательным процессом. Почва играет в жизни растения роль среды укрепления, водоснабжения и минерального питания.
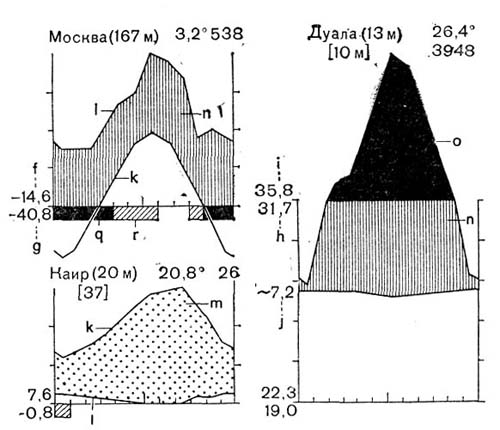
Рис.57. Климадиаграммы Москвы, Каира и Дуалы. Двенадцать делений горизонтальной шкалы соответствуют одному году. Месяцы с годовой температурой ниже 0° закрашены на базовой полосе черным; месяцы, в которые возможны заморозки и морозы, заштрихованы. Кривые отражают годовой ход температур и осадков. Одному делению вертикальной шкалы температур соответствует 5°С, а одному делению шкалы осадков 10 мм. При таком соотношении масштабов засушливой части года соответствуют месяцы, когда температурная кривая располагается выше кривой осадков. Обратное расположение выделяет влажную часть года. Указаны также высота над уровнем моря, средняя годовая t°G, годовое количество осадков, число наблюдений. Наиболее важные обозначения: I — кривая среднемесячного количества осадков; h — кривая среднемесячных температур; m — засушливый период; п — влажное время года; о — среднемесячное количество осадков, превышающее 100 мм (уменьшено в 10 раз).
Оценивая почву как среду укрепления, надо учитывать, что для укрепления в почве растение должно проникнуть в нее корнями (или ризоидами) и расти в ней. С этой точки зрения слитные массы твердых горных пород, проникновение в которые возможно только по трещинам или вследствие химического разрушения породы самим растением, существенно отличаются от рыхлых песчаных наносов или измельченных продуктов длительного физического и химического разрушения. Между этими крайностями возможны многочисленные переходы. Прочная же связь растения с почвой необходима, так как у большинства высших растений только ею обеспечивается возможность непрерывного развития особи. Отрыв от субстрата ставит растение на грань гибели.
Роль почвы в качестве среды, снабжающей растение водой, весьма велика, и различные почвы в этом отношении очень неодинаковы. Прежде всего почва должна удерживать воду. Вода атмосферных осадков, достигающая уровня грунтовых вод, присоединяется к грунтовым водам. Часть грунтовой воды под действием капиллярных сил несколько поднимается вверх. Грунтовая вода и вода капиллярной каймы доступны растению, если оно способно преодолеть корнями толщу субстрата, лежащую выше. Мощность этой толщи, очевидно, определяется глубиной залегания грунтовой воды.
Вода атмосферных осадков, выпадающая на поверхность почвы, доступна растениям в меру того, насколько она впитывается в почву и удерживается ею. Удерживаемая почвой вода может находиться в разных состояниях. Коллоидные (глинистые и гумусовые) почвенные частицы обладают отрицательным электрическим зарядом, и на их поверхности с огромной силой удерживается несколько слоев молекул воды. Эта вода прочносвязанная, или гигроскопическая. Но сила притяжения, которой обладают почвенные частицы, не исчерпывается даже при максимально возможном количестве гигроскопической воды. За счет неиспользованной силы притяжения вокруг почвенных частиц образуется более или менее толстый водный слой так называемой пленочной (рыхлосвязанной) воды. Она удерживается уже с меньшей силой, чем гигроскопическая, и отличается от нее еще и тем, что может передвигаться в жидкой фазе от частиц, где водная оболочка толще, к частицам, где она тоньше, в то время как гигроскопическая вода передвигается только при условии перехода ее в парообразное состояние. Движение пленочной воды крайне медленное, и сила тяжести на него влияния не оказывает. Прочая влага, находящаяся в почве, свободная и находится под влиянием силы тяжести и капиллярных сил. Вся прочносвязанная (гигроскопическая) вода совершенно не усваивается растением и составляет «мертвый запас» влаги. Количественно мертвый запас зависит от степени измельченности почвенных частиц. Чем они мельче, тем больше их совокупная поверхность, тем больше воды находится в прочносвязанном состоянии. Недоступность гигроскопической воды для растений объясняется огромными силами, с которыми она притягивается почвенными частицами и которые не способна преодолеть сосущая сила корней. Так, устойчивое увядание растений начинается задолго до того, как в высушиваемой почве остается только гигроскопическая вода, Из этого следует, что некоторая часть пленочной воды тоже может быть недоступной растениям.
Для оценки субстрата как среды водоснабжения надо учитывать, что часть воды, попавшей в почву, может испариться. Потеря воды при испарении в атмосферу тем больше, чем больше измельчен субстрат и чем, следовательно, сильнее развита в нем система капиллярных полостей. Но при одной и той же степени измельченности трата воды на испарение может быть различной, если в одном случае почва бесструктурна, а в другом обладает водопрочной структурой. В последнем случае между структурными частицами возникают некапиллярные промежутки, прерывающие движение воды по капиллярам по направлению к поверхности почвы. Образование структурных отдельностей обусловлено наличием в почве цементирующих веществ, прежде всего гумуса. Кроме того, возникновению структуры способствует и непосредственное участие корней. Корни, пронизывая толщу субстрата, оказывают механическое давление и уплотняют его, а сеть корней в известной мере способствует обособлению структурных частиц.
Все это очень важно для понимания взаимоотношения между растением и почвенной средой его жизни. Гумус, или перегной, представляет бесструктурную органическую массу в почве, возникающую в результате разрушения органических остатков. Если такое важное свойство, как структура почвы, создается с определенным участием веществ растительного происхождения и при активном участии корней, то не только растения зависят от почвы, но и почва зависит от растений. Кроме того, постоянное потребление воды растениями приводит к десукции почвы, дыхание корней влияет на состав почвенного воздуха, а следовательно, и на состав почвенного раствора.
К такому же выводу приводит рассмотрение почвы как источника минерального питания. Растение извлекает из почвы целый ряд элементов в значительных или крайне малых количествах (микроэлементы). Все элементы поступают в растение преимущественно в виде ионов. Ионы находятся в почвенном растворе и беспрепятственно усваиваются растением, но многие элементы находятся в почве в составе сложных соединений и усваиваются растением из твердой фазы почвы. Изучение питания растений показало, что количественное соотношение минеральных элементов в растении и в почве разное, т. е. потребление минеральных веществ растением носит избирательный характер. Минеральное питание — процесс обменный, т. е., усваивая то или иное количество катионов или анионов, растение отдает в обмен на них эквивалентное количество других ионов и катионов (чаще всего Н+ и НСО3-, которые образуются в результате выделения корнями углекислоты). Следовательно, минеральное питание растений до известной степени меняет химический состав почвы.
Кроме того, поглощенные растением минеральные вещества с опадом (отмершие листья, ветви) попадают на поверхность почвы или в самые верхние ее слои в составе более или менее сложных органических соединений. Органические соединения разрушаются гетеротрофными организмами: грибами, бактериями и др. В результате элементы минерального питания, добытые корнями, попадают на поверхность почвы и в ее верхние горизонты. Происходит, следовательно, перераспределение минеральных веществ, обогащение верхних слоев почвы, доступных корням, за счет более глубоких -происходит миграция минеральных веществ. Итак, почва как среда минерального питания с поселением на ней растительности изменяется при активном воздействии растительности.
Рельеф. Принято различать три категории рельефа — макрорельеф, мезорельеф и микрорельеф. Рельеф непосредственного влияния на жизнь растения не оказывает, но он дифференцирует поверхность и этим изменяет напряженность некоторых факторов. Следовательно, за счет рельефа увеличивается разнообразие местообитаний.
Макрорельеф создает на ограниченной площади настолько большую амплитуду высот, что в связи с этим возникает целая серия различных климатических комплексов; соответственно с высотой меняется и растительность. Вертикальная поясность горной растительности объясняется в первую очередь тем, что с высотой в горах меняется температура. Каждым 100 м подъема по вертикали соответствует снижение годовой температуры на 0,5—0,6°С. В связи с этим меняются режим осадков и условия освещения. Изменение растительности с поднятием в горы (вертикальная поясность) иногда сходна с изменением растительности на равнине по мере движения от подножия данной горной системы в сторону полюса того же полушария (горизонтальная зональность).
В горах часто приходится встречаться со сложной картиной размещения поверхностных выходов горных пород разного возраста, происхождения, физических свойств и химического состава. Они вынесены (в разных местах разные) из глубин или обнажены сопутствующим горообразовательному процессу размывом. Образовавшиеся в результате горообразования складки отделены долинами, а водные потоки, в течение больших промежутков времени бежавшие по склонам, создали сложную систему вторичных эрозионных долин, каждая из которых имеет свои склоны, уступы, водотоки. Горная местность всегда представляет массу разнообразных топологических элементов, каждый со своей экспозицией, со своими углами наклона, с различной силой размывающего действия водных потоков и с почвами разной степени выработанности, формирующимися или сформировавшимися на разных горных породах. Не удивительно, что нередко исследователь флоры встречает в горах величайшее разнообразие видов растений, в основе обусловленное пестротой жизненных условий и тем, что расселение растений в горных районах часто сдерживается высотой хребтов, через которые затруднен перенос семян. Горы часто выступают в роли настолько непреодолимых барьеров, что они становятся пограничными рубежами целых флор (Альпы и Пиренеи в Европе, Гималаи в Азии и др.).
Рельеф средних форм — мезорельеф не может иметь столь обширного и разнообразного влияния на условия жизни растений, как макрорельеф, однако он в малом масштабе повторяет картину, наблюдаемую в горах. Если разница высот в 100 м снижает температуру на 0,5 °С, то вершины холмов на равнинах (где-нибудь в предгорьях Карпат или Урала, на Донецком кряже и т. п.) могут все-таки создать «осеверенную» обстановку, пригодную для поселения леса среди степей. Северные и южные склоны, отличающиеся по инсоляции от горизонтальных плато, создают приют, соответственно, более северно и более южно распространенным растениям. Ложбины стока иной раз вскрывают водоносные пласты на склонах или пласты, своеобразные по составу породы. Со склонов в понижения стекает вода, вызывая появление вдоль водотоков более влаголюбивой растительности. Стекая со склонов, вода уносит поверхностные частицы почвы; часто наиболее плодородные откладываются в пониженных местах, создавая здесь особенно благоприятные условия минерального питания. В равнинных местностях часто на • фоне скудной растительности, покрывающей горизонтальные плато, иной раз попадаются сочлененные пышные пятна. Особенно заметны такие участки в середине лета среди разнотравных или луговых степей. И всегда такое пятно соответствует небольшому понижению, западине. Здесь создаются условия повышенного (сравнительно с соседними точками равнины) увлажнения, а дно такой западины понижено всего-то на несколько десятков сантиметров.
Чем ближе к районам, где летом бывает глубокая засуха, тем большее значение приобретают эти ничтожные понижения и повышения — элементы микрорельефа.
Рельеф создает разнообразие условий и комплексов растений в данной географической области. Благодаря рельефу фактическая площадь, пригодная для растений, возрастает.
Биотические факторы объединяют все организмы животной и растительной природы и микроорганизмы, которые могут оказать то или иное влияние на жизнь данного растения, его особей или совокупностей особей (популяций), а через это и на жизнь вида. Биотические факторы крайне разнообразны благодаря огромному видовому разнообразию организмов и специфичности жизнедеятельности каждого вида. Биотические воздействия на растения бывают прямыми (например, поедание травы животным) и непрямыми (изменение среды, в которой живет растение). Если при прямых воздействиях важно иногда воздействие одной особи животного существа на растение, то непрямое влияние становится, как правило, заметным, если оно слагается из деятельности многочисленных особей. Впрочем, прямые и непрямые влияния не всегда можно разграничить. Животные уничтожают растения (съедают, вытаптывают), выступают в роли переносчиков семян и плодов и воздействуют на растения как опылители. Влияние опылителей огромно, особенно в случаях, когда цветки растения приспособлены к опылению только одним опылителем или очень ограниченным кругом их.
Большую роль в жизни растений играет животный мир почвы. Животные подгрызают корни, размельчают и переваривают остатки растений, передвигаются в почве, разрыхляя ее, пропуская через кишечник частицы почвы, изменяют их химизм и строение. Земляные муравьи и землерои из числа позвоночных (кроты, суслики, мышевидные грызуны) насыпают холмики на поверхности почвы, создавая условия для поселения сорных, растений.
При массовом распространении роющих животных они превращают всю площадь какого-нибудь сообщества (луга, степи и т. п.) в неустойчивую среду, где свежие холмики из выбросов на поверхность материалов более глубоких почвенных горизонтов чередуются с соседними участками, еще не покрытыми выбросами. Холмики, насыпанные роющими животными, отличаются от соседних, еще не засыпанных площадей в химическом отношении, особенно если выносимый на поверхность материал извлекается из заметной глубины. Тогда может возникнуть местное засоление или, наоборот, опреснение субстрата. Отличаются такие холмики и в физическом отношении — плотностью сложения, размерами и прочностью структуры. Кроме того, здесь устанавливается особый режим водоснабжения. Выступая над общим уровнем поверхности, они легче иссушаются. В целом поверхностные образования такого рода образуют отличную от соседних точек среду жизни растений — благоприятную для одних организмов и мешающую развитию других. С течением времени, по мере того как возникают новые сурчины и сусликовины, старые размываются дождями и разрушаются ветрами. Атмосферные воды вымывают из них растворимые соединения, вновь унося их в глубины почвы. Через некоторое время эдафиче-ские условия жизни на месте прежних выбросов сравниваются с окружающей ровной площадью.
На этом примере наглядно видно, что деятельность животных может дифференцировать, разнообразить даже такой, казалось бы, консервативный элемент среды растительной жизни, как почва,— продукт очень длительного исторического изменения поверхностных выходов геологических напластований под совместным влиянием климата и организмов.
Влияние растений на другие растения также разнообразно. Отметим прежде всего влияние паразитов. Особенно многочисленны они среди грибов. Таковы ржавчинные, головневые грибы и многие другие, способные быстро распространяться и охватывать своим вредоносным действием огромные количества особей растения-хозяина.
Паразиты представляют такую форму односторонне выгодного сожительства, когда между растениями устанавливается принужденный обмен веществ. Случаи, когда хозяин способен не только оказывать сопротивление паразиту, но и извлекать из него некоторую пользу, послужили, вероятно, основой выработки в процессе эволюции особой формы сожительства (взаимопаразитизма), причем достаточно устойчивой и производящей впечатление смирного сожительства». Такое сожительство называется симбиозом. Примерами могут служить микоризы, лишайниковые симбиозы.
Сожительство растений может быть и без прижизненного обмена веществ. В этих случаях растение, живущее на другом, используя последнее только как место прикрепления, называют э п и-ф и т о м. Частным случаем эпифитизма являются эпифиллы, т. е. растения, использующие в качестве опоры только листья другого растения. Эпифиты и эпифиллы могут заметно влиять на свой субстрат, затрудняя газообмен и другими путями.
Сходную по отсутствию прижизненного обмена веществ категорию представляют лианы-растения, имеющие недостаточно твердые и устойчивые стволы и ветви достаточно высоких растений или обвиваются вокруг их стволов, порою мешая росту последних в толщину («душители деревьев»).
Но чаще всего влияние фитобиотического фактора проявляется в форме изменения среды данного растения жизнедеятельностью окружающих его прочих растений. В основном это влияние видно в изменении световых условий, уменьшении нагрева, препятствии ночному охлаждению, в задержании части осадков, повышении влажности воздуха, иссушении почвы корнями, а также в конкуренции за элементы минерального питания и за воду.



